Introduction
Mechanotransduction is essential for sensing touch, hearing, and autonomic functions such as blood pressure regulation, gastrointestinal (GI) motility, cell differentiation and development, muscle stretch, and vascular tone (, ). In a two-step process, cells first sense mechanical stimuli through ion channels, transmembrane adhesion receptors, sarcomeric proteins, and cell surface receptors (–). These mechanoreceptors then convert mechanical forces into electrochemical signals that trigger appropriate cellular responses. However, defects in mechanotransduction are found in various genetic and acquired diseases, ranging from muscular dystrophy, cancer, and cardiomyopathies to GI disorders (, , ). Highly specialized electromechanical organs, like the heart and GI tract, generate tissue-specific mechanical forces sensed by specialized mechanosensitive cells (, ). The layers of the GI tract, including the mucosa, submucosa, and muscularis mucosae, have distinct mechanical properties that depend on specialized mechanoreceptor cell types. Coordination of these mechanical activities is essential for the peristaltic reflex, segmentation, and migratory motor activity that facilitate the digestion, absorption, and propulsion of food. In the GI epithelium, distinct sensory cells, like enterochromaffin cells, secrete serotonin (also known as 5-hydroxytryptamine [5-HT]) in response to epithelial mechanical forces, and thereby regulate intestinal fluid secretion (). Neuronal cells in the GI tract, like intrinsic primary afferent neurons and extrinsic sensory neurons, sense physical forces and convert them into electrical signals (). Nonsensory cells like interstitial cells of Cajal, smooth muscle cells, and immune cells respond to physical stimuli by altering the inherent functions specific to each cell type, be it motility, synthesis, or secretion (, ). Thus, responding to mechanical forces is an inherent and fundamental process for normal GI functions. However, disturbed mechanotransduction in the GI tract has been implicated in diverticulosis, intestinal pseudo-obstruction (), chronic constipation (), visceral hypersensitivity (, ), functional dyspepsia (), irritable bowel syndrome, colon cancer (), inflammatory bowel disease (), and even systemic conditions, like obesity ().
The initial step in mechanotransduction involves the activation of cell surface receptors or ion channels. Cells termed mechanoreceptors that transmit their mechanical cues to sensory neurons are deemed mechanosensitive. Several mechanosensitive ion channels, such as L-type voltage-gated calcium channels (e.g., Cav1.2), voltage-gated sodium channels (e.g., Nav1.5), Ca2+-activated large-conductance potassium channels (e.g., BKCa), and nonselective cation channels (e.g., TRPV2, TRPV4, TRPC4, TRPC6, TRPC7), regulate GI motor function and secretion and are associated with motility disorders (, ). Mechano-gated TREK1 channels expressed in the smooth muscle in the ileum and colon and TRAAK channels in enteric neurons modulate gut motility (, ). In contrast to the GI tract, the mechanosensing properties of liver cells have not been studied extensively, partly because of the lack of sensory innervation within the liver parenchyma. However, abnormal physical forces within the liver influence the phenotype of hepatic stellate cells, hepatocytes, portal fibroblasts, and endothelial cells and, in turn, contribute to hepatic fibrosis and cancer (–).
Although it has been underappreciated, the pancreas is a highly mechanosensitive organ that is susceptible to mechanical injury (–). Slight manipulations of the pancreas during abdominal surgery or abdominal trauma may induce pancreatitis, and animal models demonstrated that activation of mechanosensors on pancreatic acinar cells was the root cause (–). Pancreatic stellate cells also express mechanically activated ion channels that are coupled to cell activation and pancreatic fibrosis (, , ). Surprisingly, pancreatic β cells possess mechanically activated ion channels that appear to be coupled to insulin secretion (, ).
Insights into the mechanism of mechanosensing have been revealed by the recent discovery of a class of evolutionarily conserved mechano-gated ion channels known as Piezo (). Piezo channels are a family of mechanically activated ion channels consisting of Piezo1 and Piezo2. Piezo1 is a large cation channel that is highly permeable to Ca2+ (, , ) and is activated by mechanical forces including static pressure, fluid shear stress, and stretch (, , , , ). It is expressed primarily in nonsensory tissues including the vascular and lymphatic systems, lung, bladder, skin, GI tract, and pancreas (, , ). Piezo2 is typically expressed in tissues that respond to physical touch, including sensory neurons. The molecular structure of Piezo channels has been determined by cryo–electron microscopy. The predicted size of approximately 2,500 amino acids is composed of a three-bladed propeller-shaped homotrimeric structure, 38-transmembrane regions, and a central pore. Piezo1 and Piezo2 share nearly 42% sequence homology (, , ). Both Piezo1 and Piezo2 are fast-activating and inactivating channels (, –). Endogenous activators of Piezo channels have yet to be identified, but several studies suggest that alteration of membrane lipids could modulate Piezo1 gating properties (). It is possible that cholesterol could modulate Piezo1 and Piezo2 channel activities via cholesterol-binding stomatin-like protein 3 (STOML3), which physically interacts with the channels (). STOML3 oligomerization reversibly reduces the sensitivity of Piezo currents in sensory neurons and tunes Piezo channel sensitivity at molecular-scale stimuli relevant to delicate touch. Moreover, Piezo1 gating properties depend on the cell’s intrinsic fatty acid composition (, , ). Matrix stiffness can regulate the sensitivity and degree of activation of mechanical ion channels including Piezo1 (–). Specifically, the collagen IV network of extracellular matrix (ECM) increases the sensitivity of Piezo1 to mechanical stimuli ().
Several gain-of-function and loss-of-function mutations in Piezos have been linked with various hereditary human diseases, including hereditary xerocytosis, congenital lymphatic dysplasia, and an autosomal recessive syndrome of muscular atrophy with perinatal respiratory distress. Altered Piezo channel signaling has been associated with a variety of physiological and pathophysiological processes, including cell division, axon growth and regeneration, erythrocyte volume regulation, vascular hyperpermeability (), chronic obstructive pulmonary disease, and pressure-induced pancreatic and renal (, ) fibrosis, touch, proprioception, pain, baroreceptor reflex regulation, urinary function, and innate immunity (, , , –). In addition, Piezo channels play an important role in tumor development and cancer cell metastasis (, –).
Piezo channels are expressed in many cells of the GI tract and are essential in proliferation, differentiation, secretion, and motility. This Review focuses on our current understanding of Piezos in GI function and disease. Studies using cell- or tissue-specific Piezo channel gene deletion in mice, siRNA-mediated gene silencing of Piezo channels, or pharmacological blocking of Piezo channels in vitro indicate that Piezo channels have a crucial role in GI cancer, fibrosis, inflammation, pain, secretion, and immunity.
Mechanotransduction in pancreatic disease
Physical manipulation of the pancreas by surgery or endoscopic pancreatography may induce pancreatitis. The latter condition can be modeled experimentally by artificial increasing of pancreatic duct pressure (). In the process of isolating pancreatic acinar cells, we noticed that mechanically disrupting the pancreas activated calcium-sensitive 5-lipoxygenase (). Positing that this must be the result of increased intracellular calcium ([Ca2+]i), we discovered that Piezo1, an inherently mechanosensitive channel, is abundantly expressed in the pancreas (, ). Two approaches were used to assess the mechanosensitivity of pancreatic acinar cells. First, using a micropipette, mechanical force was applied to the surface of acinar cells loaded with a calcium-sensitive dye (). Not only did touching a cell produce a rapid increase in [Ca2+]i, but the calcium signal was relayed to adjacent cells in a sequential manner consistent with the unit functionality of an acinus (). Screening a library of 3 million compounds, Patapoutian’s group identified a selective Piezo1 agonist, Yoda1 (). When applied to pancreatic acini, Yoda1 induced an increase in [Ca2+]i, reproducing the effects of mechanical force. Thus, Piezo1 confers mechanical sensitivity to pancreatic acinar cells through its calcium channel properties.
Acute pancreatitis.
In pancreatic acinar cells, prolonged elevation in intracellular calcium concentration ([Ca2+]i) disrupts mitochondrial function, reduces ATP synthesis, and initiates zymogen granule–lysosome fusion and intracellular enzyme activation leading to pancreatitis (, ). Therefore, for self-preservation, acinar cells possess multiple safeguards to ensure [Ca2+]i homeostasis. Once Piezo1 was discovered in pancreatic acini, it seemed possible that excess mechanical stimulation could cause pancreatitis through a calcium-sensitive mechanism (Figure 1), implying that precise regulation of [Ca2+]i was necessary to prevent pancreatitis.
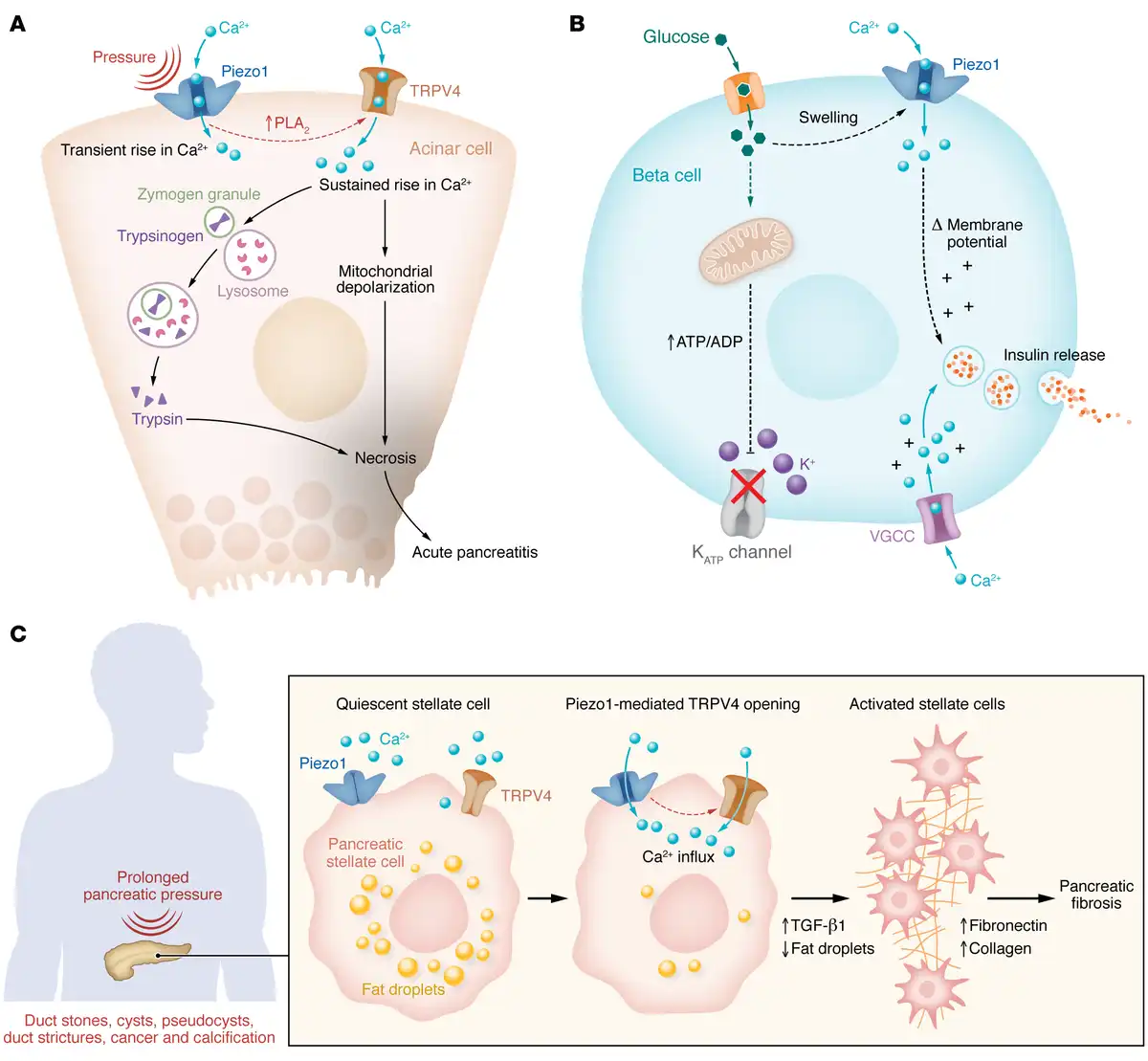
Figure 1
Model of mechanosensing in the pancreas.
(A) Piezo1-induced TRPV4 opening causes intracellular calcium ([Ca2+]i) elevation in acinar cells and is responsible for pressure-induced pancreatitis. Pressure or shear stress force opens Piezo1 channels, which induces phospholipase A2 (PLA2) and activates TRPV4. Sustained elevation in [Ca2+]i leads to mitochondrial depolarization, lysosome–zymogen granule fusion, trypsinogen activation, and pancreatitis. (B) Piezo1-induced [Ca2+]i elevation through glucose-triggered dynamic changes in volume and deformation in cell membrane facilitates insulin release from pancreatic β cells. (C) Pressure-sensing Piezo1 channel signaling activates PSCs. The activated phenotype lacks perinuclear fat droplets and shows increased levels of TGF-β1, fibronectin, and type I collagen that contribute to pancreatic fibrosis. VGCC, voltage-gated calcium channel.
To evaluate this possibility, pressure-induced pancreatitis was produced in mice via pancreatic duct injection, a model of clinical pancreatitis that occurs as a complication following endoscopic retrograde cholangiopancreatography (ERCP) (). Pancreatitis was blocked by treatment of mice with the Piezo1 inhibitor GsMTx4 (, ). Global deletion of Piezo1 is embryonic lethal because of the wide distribution of Piezo1 in many tissues, including vascular endothelium, but conditional deletion of Piezo1 in pancreatic acinar cells did not cause any detectable defect (, ). Importantly, mice with acinar cell–selective Piezo1 knockout were protected against pressure-induced pancreatitis. The Piezo1 agonist Yoda1 increased [Ca2+]i in pancreatic acinar cells and, in high concentrations, caused a sustained [Ca2+]i elevation that led to sustained mitochondrial depolarization, trypsinogen activation, and lactate dehydrogenase (LDH) release, reproducing many features of pancreatitis in vitro (, ) (Figure 1A).
Since Piezo1 is a rapidly inactivating channel, Piezo1 opening produces only transient elevation in [Ca2+]i that alone was insufficient to cause pancreatitis. Therefore, it was not known how pressure produced the prolonged [Ca2+]i elevation necessary to induce pancreatitis. A series of in vitro and in vivo studies showed that the amount and duration of pressure applied to the pancreas determined the level and duration of [Ca2+]i elevation in acinar cells and corresponded to pathological changes, e.g., intracellular trypsinogen activation and LDH release (). Moreover, Piezo1 activation, caused by high pressure of long duration, triggered transient receptor potential vanilloid 4 (TRPV4) channel opening by activating phospholipase A2 (PLA2) and stimulating the conversion of arachidonic acid to epoxyeicosatrienoic acid, an endogenous activator of TRPV4 channels. In other words, the pathological consequences of pressure required TRPV4 activation (, , ) (Figure 1). Pressure-induced pancreatitis was reduced in mice with either acinar cell–specific deletion of Piezo1 or global deletion of Trpv4. Thus, pressure-induced pancreatitis is caused by Piezo-initiated calcium signaling that requires TRPV4 to become fully manifest ().
Chronic pancreatitis and fibrosis.
Chronic pancreatitis is caused by persistent or repetitive injury to the pancreas and is characterized pathologically by gland atrophy, inflammatory cell infiltration, and fibrosis (–). Pancreatic fibrosis consists largely of collagen, fibronectin, and other ECM proteins that are produced and secreted by activated pancreatic stellate cells (PSCs) (–). Pancreatic fibrosis also develops under conditions of elevated pancreatic pressure, such as prolonged pancreatic duct obstruction or cancer, raising the possibility that pressure induces fibrosis and that PSCs are pressure sensitive (, , , , ). Consistent with this idea, PSCs were found to express Piezo1 (). In a [Ca2+]i-dependent manner, shear stress or the Piezo1 agonist Yoda1 transformed PSCs from a quiescent to an activated phenotype characterized by loss of perinuclear fat droplets and increased TGF-β1, fibronectin, and type I collagen expression () (Figure 1). Like acinar cells, human and mouse PSCs also express TRPV4. Although TRPV4 does not have direct mechanosensing properties, in PSCs it is linked to Piezo1; and mice with either stellate cell–specific deletion of Piezo1 or global Trpv4 deletion were protected against pressure-induced pancreatic duct fibrosis (). Thus, mechanical forces directly activate PSCs and induce pancreatic fibrosis.
Piezo1-mediated endothelial dysfunction.
One-quarter of pancreatitis patients develop vascular complications, which contribute to the high mortality of severe pancreatitis (, ). Vascular hyperpermeability is a critical feature that promotes edema and vascular collapse, and severe pancreatitis can be complicated by splanchnic venous system thrombosis (, ). Piezo1 is highly expressed in vascular endothelium and may be activated by vascular shear stress or elevated intravascular pressure, leading to endothelial dysfunction (, ). We recently reported that Piezo1 mediates pathological endothelial cell responses to high-shear stress (). Activation of Piezo1 by prolonged shear stress in endothelial cells produced a sustained elevation in [Ca2+]i and increased PLA2 activity, which in turn caused TRPV4 channel opening, leading to cytoskeletal disorganization, increased expression of the adhesion protein VCAM1, and loss of adherens junctions. This disassembly of adherens junctions and elevated VCAM1 promoted leukocyte adhesion (). These findings suggest a mechanism by which pathological Piezo1 activation within the endothelium may contribute to the vascular complications and edema that accompany acute pancreatitis.
Piezo regulation of β cell function.
Glucose is the major physiological stimulant of insulin secretion from pancreatic β cells. Glucose and hypotonicity also induce cell swelling. Piezo1 was recently identified in β cells (), and through a [Ca2+]i-dependent mechanism Piezo1 activation was shown to stimulate insulin secretion in β cell lines and isolated mouse islets () (Figure 1). Although the extent to which cell swelling contributes to insulin secretion in vivo is unknown, the observation that β cell–specific Piezo1-knockout mice exhibited impaired glucose tolerance suggests that Piezo1 may have a physiological role in insulin secretion. Interestingly, Piezo1 expression is elevated in islets from humans with type 2 diabetes and in the db/db mouse model of diabetes, in which Piezo1 translocates from the plasma membrane to the nucleus (). Translocation of Piezo1 eliminates Piezo1-induced Ca2+ influx, and insulin secretion is reduced.
Mechanosensing in the gut
GI motility is critical for the ingestion, propulsion, and digestion of food. The GI tract is composed of several types of mechanosensitive cells, including enterochromaffin cells of the mucosa, enteric neurons, smooth muscle cells (SMCs), and interstitial cells of Cajal (). These cells express an array of voltage-gated calcium and sodium channels, potassium channels, and nonselective cationic channels that appear to respond to mechanical forces but lack inherent mechanosensing properties intrinsic to the channel itself. For this discussion, we refer you to several comprehensive recently published reviews on these channels (, , , , , , –).
Piezo channels are expressed in the epithelia, enteric nervous system, and SMCs of the digestive system as well as throughout the vascular smooth muscle and endothelium in humans and mice (, , , , , ). Gd3+, a nonspecific mechanical ion channel blocker, inhibits Piezo-type current in SMCs of the GI tract, suggesting a potential role of Piezo1 channels in peristalsis and other stretch-related functions in the gut (, ). Little is known about Piezo in myenteric plexus and intestinal muscularis cells (). In the intestine, most epithelial cells, including goblet cells, Paneth cells, enterocytes, and endocrine cells, respond to mechanical forces, including stretch/distension, membrane distortion and deformation, shear stress, touch, tensile force compression, intraluminal pressure, and cell volume changes (, , –). Static forces in the gut are essential for epithelial cell proliferation, differentiation, and turnover (). Piezo1 is present in most epithelial cells in the gut and can regulate epithelial cell proliferation and differentiation (, ). In the Drosophila gut, Piezo1 triggers the differentiation of stem cell progeny (). In addition, Piezo1 controls normal epithelial homeostasis by inducing the extrusion of live cells that maintain the number of cells in the epithelia during proliferation-induced overcrowding (). In contrast to Piezo1, Piezo2 is expressed in sensory neurons and primarily functions as a nociceptor (, , , –). However, it also regulates light touch () and is an important sensor in the GI tract.
Piezo regulation of food intake.
External sensory cues and internal metabolic states control animal feeding. In Drosophila, Piezo-expressing neurons innervating the crop (equivalent to the mammalian stomach) sense mechanical cues of stomach fullness and prevent food overconsumption (, ). These neurons reside in the pars intercerebralis, a neurosecretory center in the brain, and express insulin-like peptides that regulate food intake and metabolism. Piezo knockdown led wild-type flies to eat more and exhibit an overconsumption phenotype similar to that of piezo-null mutant flies. Expression of either mammalian Piezo1 or fly piezo in these neurons of piezo-null mutants prevented the overconsumption phenotype ().
Nutrient sensors in the gut identify food rich in nutrients. A subset of piezo-expressing neurons also express diuretic hormone 44 (DH44), which helps flies detect sugar and operate during starvation. Piezo knockdown in DH44 neurons, which also project to the Drosophila crop, stimulates DH44 neuronal activity and food intake in fed flies (). Together these observations indicate a role for Piezo channels in the regulation of satiety and food intake and open similar possibilities in mammals.
Piezo2 in esophageal peristalsis.
Esophageal peristaltic movements are controlled by reflexes executed by vagal motor nuclei in the hindbrain, nucleus ambiguus, and esophageal enteric ganglia (, ). Clinically, esophageal dysmotility is typified by dysphagia, although the etiologies are diverse (). The esophageal epithelium is innervated by several types of mechanosensory neurons with distinct electrophysiological properties and region-specific nerve endings such as intramuscular arrays, mucosal endings, and intraganglionic laminar endings (). Single-cell RNA sequencing analysis identified two transcription factors, Prox2 and Runx3, that distinguish most vagal nodose neurons positive for Piezo2 (). Three subtypes of Prox2 and Runx3 vagal neurons are low-threshold mechanoreceptors that innervate the esophagus and stomach, forming intraganglionic laminar endings on the enteric ganglia. Ablation of Prox2 and Runx3 neurons using intersectional genetic tools, followed by videofluoroscopic swallowing studies, in freely behaving rodents showed that esophageal motility requires these neurons (). Esophageal intramuscular array neurons positive for Piezo2 also respond to stretch. It appears that Piezo2-positive vagal neurons respond to mechanical signals in the esophagus and control peristalsis, raising the possibility that they could have a role in dysphagia.
Piezo in gut motility, secretion, and visceral sensitivity.
Enterochromaffin cells are a subtype of enteroendocrine cells and the primary source of serotonin. It has long been known that mechanical forces release serotonin, which stimulates GI motility and activates serotonin-sensitive colonic afferent neurons (). However, it was not until the discovery of Piezo2 in enterochromaffin cells that it was appreciated that these effects were mediated through cell surface mechanically activated ion channels (, ). It was recently shown that applying a Piezo2 blocker or genetic deletion of Piezo2 in enterochromaffin cells reduced stretch-induced Ca2+ signaling, serotonin release, and intestinal secretion (, ).
Irritable bowel syndrome (IBS) is a constellation of abdominal pain and alterations in bowel motility with diarrhea, constipation, or both. The underlying cause is unknown, although heightened visceral nociception is a prominent feature in many patients. Enterochromaffin cells communicate with enteric nerves through paracrine or synaptic connections comprising a semiautonomous effector network coupled to the parasympathetic and sympathetic nervous systems (, ). This bidirectional gut-brain connection involving serotonergic pathways influences both intestinal and extraintestinal symptoms in IBS. Elevated levels of serotonin have been reported in IBS, and 5-HT3 receptor antagonists and 5-HT4 receptor agonists have been used to treat either diarrhea- or constipation-predominant IBS, respectively, indicating that serotonin is involved in the pathogenesis of IBS (–).
Piezo2 in enterochromaffin cells might be linked to the pathophysiology of IBS. Stretching or elevated pressure within the GI tract activates Piezo2 in enterochromaffin cells, which release serotonin; therefore, serotonin-induced symptoms could arise from either excessive force producing hyperalgesia, or increased sensitivity to normal force (allodynia) (). It remains to be determined whether conditions such as prolonged pressure in IBS can alter Piezo2 expression and influence downstream serotonin signaling. Both Piezo1 and Piezo2 are expressed in the enteric nervous system of mice, guinea pigs, and humans (). However, Piezo1 is absent in sensory afferents. In contrast, Piezo2 is extremely rare in enteric neural somata but is enriched in dorsal root ganglion (DRG) neurons (which receive sensory and nociceptive input and transmit it to nerves ascending to the CNS), including small-diameter unmyelinated neurons, suggesting a possible role in mechanonociception (). Growing evidence suggests that Piezo2 is critical for visceral hypersensitivity. In a rat model, neonatal colonic instillation of acetic acid induced visceral hypersensitivity, which was blocked by Piezo2 knockdown in DRG neurons, indicating that Piezo2 mediated the hypersensitivity response (). Loss-of-function mutation in Piezo2 protects against pain sensitization. In contrast, activating Piezo2 in sensory neurons induces noxious stimuli (, , ).
In a classic study in IBS patients, heightened nociception was demonstrated in response to colonic balloon distention (). Visceral pain is transmitted to the spinal cord via visceral afferent neurons, a subtype of which express the ion channel TRPV1. TRPV1 has been implicated in pain originating in many tissues, including the intestine, pancreas, and bladder (). Mice with genetic deletion of TRPV1 exhibited reduced sensitivity to colorectal distension (–). Interestingly, Piezo2 is expressed in TRPV1-lineage nociceptors in the colon (). Ablating either Trpv1 or Piezo2 in TRPV1-expressing colonic neurons reduced action potential firing by visceral afferents and visceromotor response in mouse models of zymosan-induced IBS and partial colon obstruction, raising the possibility that Piezo2 and TRPV1 could be targets for treating visceral pain caused by mechanical distension or stretch and may be relevant to visceral pain conditions such as inflammatory bowel disease, IBS, or intestinal obstruction () in which either excessive stretch or hypersensitivity to normal stretch may occur (Figure 2).
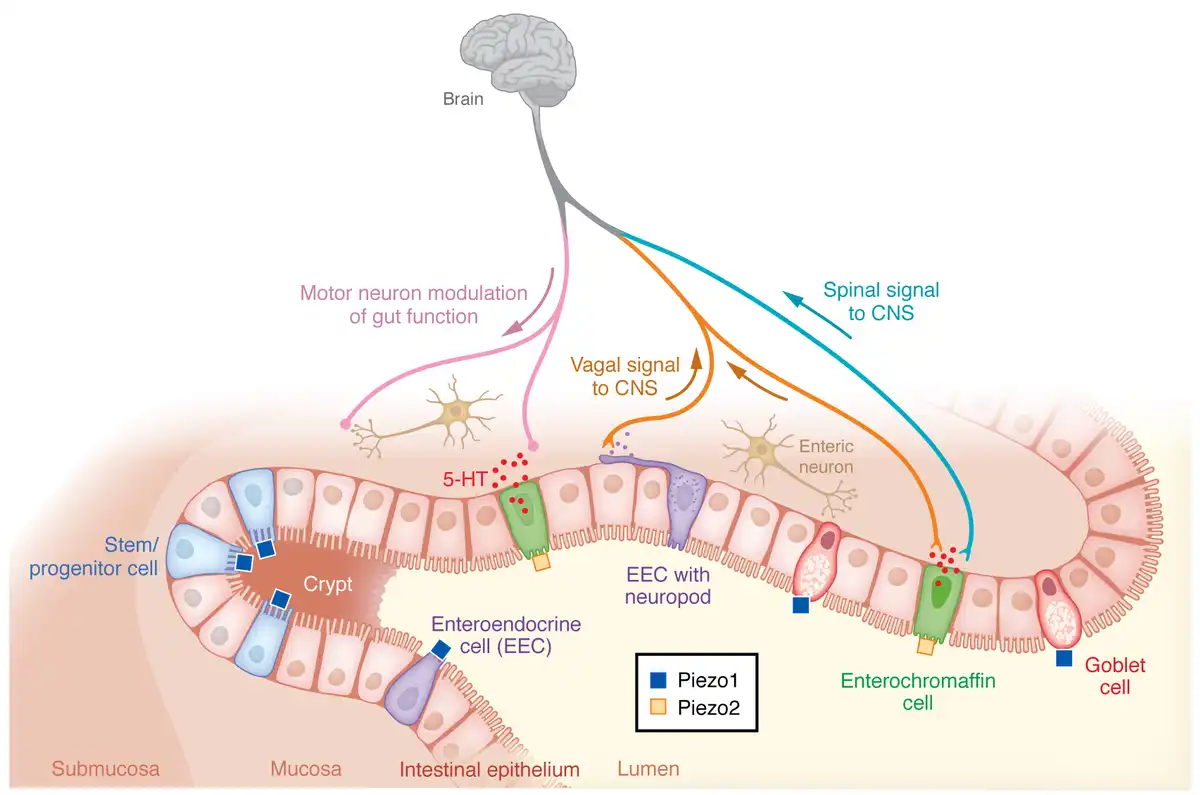
Figure 2
Schematic depiction of mechanosensing functions of Piezo1 and Piezo2 in the gut.
Piezo1 is expressed on stem cells of the intestinal crypt, enteroendocrine cells (EECs), and goblet cells and stimulates stem cell proliferation, EEC differentiation, and mucus production, respectively. Piezo2 in enterochromaffin cells triggers 5-HT release and mediates visceral hypersensitivity.
Piezo1 in GI cancer.
Mechanical forces are essential for normal epithelial function. In contrast, abnormalities in force or altered sensation can lead to disease, including cancer (, , ). A unique characteristic of Piezo1 in the GI epithelium is its ability to sense cell crowding () and regulate cell migration (). In GI epithelium, tumor growth exerts pressure on the adjacent epithelium. This altered mechanical stress induces aberrant crypt foci (), suggesting that stem cell differentiation and cell migration are sensitive to surrounding forces (, , ).
Piezo1 channels are expressed in squamous cells of the esophageal mucosa, and recent studies demonstrated that Piezo1 levels are elevated in human esophageal squamous cell carcinoma (ESCC) tumors (, ). Mechanistically, Piezo1 promotes cancer cell metastasis and invasion by promoting cell migration, elevating the production of angiogenic factors, and enhancing matrix remodeling (, , , , , , ). These responses appear to be direct cellular effects of Piezo1, since downregulation of Piezo1 using shRNA inhibited the proliferation, migration, and invasion of ESCC cell lines EC109 and EC9706. Piezo1 downregulation inhibited epithelial-mesenchymal transition and restored the epithelial cell phenotype (). Recent studies also indicate that Piezo1 plays a key role in cancer evolution. Downregulation of Piezo1 induced apoptosis via the Piezo1/p53/Bax/caspase-3 axis and inhibited G0/G1-to-S-phase cell cycle progression (). In addition, Piezo1 monoclonal antibodies that induced Piezo1 internalization in human ESCC tumor cells when administered together with the antineoplastic agent monomethyl auristatin E preferentially killed ESCC tumor cells with high Piezo1 expression and suppressed tumor progression in ESCC xenograft tumor models, suggesting that Piezo1 may provide a novel target for ESCC ().
Mechanical forces within tumors themselves regulate Piezo1 signaling that affects tumor growth and metastasis; and clinically, upregulation of Piezo1 expression in colon cancer is associated with a poor patient prognosis (, ). It appears that mechanical stress triggers Piezo1 signaling, leading to reduced expression of mitochondrial calcium uniporter, increased expression of HIF-1α and VEGF, and decreased mitochondrial membrane potential production, which promote colon cancer cell metastasis (, , ). Knockdown of Piezo1 channels by siRNA blocked the effects of mechanical stress to promote tumor malignancy and cancer metastasis in human colon cancer cell lines (HCT-116, SW-480) and similarly promoted gastric cancer in a xenograft model of human gastric tumors in BALB/c nude mice (, ).
Immune cell actions of Piezo1.
Piezo1 has been linked to inflammation and immune surveillance in cancer (, , , –). Piezo1 is highly expressed in myeloid cells, where it is the primary mechanosensor linking mechanical stress to immune regulation. This function appears to be relevant to cancer surveillance, as deletion of Piezo1 in myeloid cells reduced pancreatic cancer progression (, ). Piezo1 is essential to the immune response in dendritic cells (, ). In a syngeneic ovalbumin mouse model, mechanical force upregulated production of the proinflammatory cytokines IL-6 and TNF-α in a Piezo1-dependent manner, and Piezo1-deficient dendritic cells suppressed the antitumor response ().
In addition to dendritic cells, tumor-associated macrophages (TAMs) play a substantial role in cancer-related inflammation and are critical regulators of tumorigenesis, including gastric cancer (, ). Like many cancers, the gastric and colorectal tumor microenvironments harbor myeloid-derived suppressor cells, regulatory T cells, and TAMs and support tumor growth and progression (–). TAMs are derived from chemokine receptor type 2 inflammatory monocytes and consist of two main types, M1 and M2. M2 macrophages promote tumor progression by (a) triggering tumor neovascularization through the secretion of pro-angiogenic and inflammatory factors and (b) generating an immunosuppressive microenvironment. TAMs also can enhance tumor cell invasion and migration by promoting epithelial-mesenchymal transition and ECM remodeling (). Furthermore, TAMs can reduce the effectiveness of cancer treatments, including chemotherapy, and response to immune checkpoint inhibitors and radiotherapy (). Importantly, Piezo1 is necessary for macrophage polarization, raising the possibility that Piezo1 blockade might prevent the permissive effects of M2 TAMs (, ). The observation that genetic deletion of Piezo1 in myeloid cells protected against cancer suggests that Piezo1 could be a target for suppressing inflammation in different organs, including GI tract cancers (), although the potential for immunosuppressive side effects of such a strategy is unknown.
Piezo1 in Crohn’s disease.
Crohn’s disease is a chronic intestinal inflammatory disease that can affect any region of the GI tract but most commonly involves the ileum and proximal colon. Initiation of proinflammatory responses in the intestine likely involves Piezo1, which is highly expressed in the ileum of patients with Crohn’s disease and positively correlates with the Crohn’s Disease Activity Index and fecal calprotectin levels (). Activation of CD4+ T cells is a key feature in inflammatory bowel disease, and loss of Piezo1 in CD4+ T cells promotes Th1 and Th17 cell polarization. The subsequent reduction in inflammatory signals indicates that Piezo1 regulates the inflammatory response of pathogenic T cells. In a mouse model of chronic colitis, Piezo1 in CD4+ T cells was critical for development of intestinal inflammation (), raising the possibility that Piezo1 activation could promote intestinal inflammation.
Piezo1 in goblet cells.
Intestinal mucus is produced by goblet cells and limits the exposure of epithelial cells to bacteria. Defects in the gut mucus layer allow pathogenic bacteria to reach the epithelium and promote intestinal inflammation. Piezo1 is expressed in goblet cells and regulates mucus synthesis and secretion. Mice with goblet cell–specific Piezo1 knockout exhibited thinning of the intestinal mucus layer, elevated inflammatory cytokines (e.g., CXCL1, CXCL2, IL-6), and an increased number of pathogenic bacteria in the gut ().
In humans, functional gut disorders are often linked with chronic stressful life events (–). Water avoidance stress has been used to model psychological stress in mice and induces colonic epithelial barrier dysfunction, colorectal hypersensitivity, and dysbiosis. It was recently shown in mice that water avoidance stress diminished Piezo1 expression in goblet cells, reduced mucus barrier function, and impaired intestinal motility (). The Piezo1 agonist Yoda1 improved mucus barrier function and reversed the defect in intestinal motility. Although these studies point to a beneficial effect of Piezo1 on intestinal barrier function by maintaining the mucus layer, there is evidence that Piezo1 may also negatively regulate epithelial tight junction function (). Overexpression of Piezo1 or stimulation of Piezo1 by Yoda1 decreased the tight junction protein claudin-1, leading to impaired colon epithelial integrity and loss of barrier function.
Overall, it appears that excessive Piezo1 activity has deleterious effects on intestinal immunity and epithelial tight junctions. Whether its positive effects maintaining intestinal mucus production outweigh these other actions remains to be determined.
Conclusions
Mechanically activated ion channels detect physiological pressure, shear force, stretch, and stiffness, and promote normal biological responses including cell growth, migration, turnover, and secretion. Abnormally high or prolonged mechanical forces induce pathological changes that are manifested by cellular and tissue damage that initiates an inflammatory response. In the pancreas, acute pancreatitis occurs when acinar cells are subjected to excessive force, and fibrosis follows chronic stellate cell stimulation. The observation that Piezo1 is required for normal insulin secretion raises the intriguing possibility that mechanosensing may be a common feature of exocytosis in other cell types. Piezo channels in enterochromaffin cells link pressure and distension in the gut to the release of serotonin and its pathological consequences in IBS. By virtue of its expression in immune cells, Piezo1 contributes to intestinal inflammation and immune surveillance. Piezo channels are upregulated in GI cancers including gastric cancer and colorectal carcinoma, and, by virtue of their ability to sense cell crowding, these receptors may be involved in cancer metastasis. Despite the attraction of mechanoreceptor blockade as a possibility for treating or preventing inflammation or cancer, the wide distribution of Piezo channels in non-target tissues may limit the utility of Piezo-targeted pharmacotherapies. Perhaps development of tissue-selective therapies or therapies directed at pathways downstream of Piezo signaling would mitigate potential off-target effects.
This work was supported by NIH grants DK124474, DK120555, DK125308, and 10BX006301 from the Department of Veterans Affairs, and National Pancreatic Foundation Research Grant 2022. The authors thank Steve Vigna for helpful advice and for reviewing the manuscript.
- 1. Douguet D, Honoré E. Mammalian mechanoelectrical transduction: structure and function of force-gated ion channels Cell. 2019;179(2):340–354
- 2. Ranade Sanjeev S, et al Mechanically activated ion channels Neuron. 2015;87(6):1162–1179
- 3. Gayer CP, Basson MD. The effects of mechanical forces on intestinal physiology and pathology Cell Signal. 2009;21(8):1237–1244
- 4. Liu Y, et al Molecular tension probes for imaging forces at the cell surface Acc Chem Res. 2017;50(12):2915–2924
- 5. Wolfenson H, et al Steps in mechanotransduction pathways that control cell morphology Annu Rev Physiol. 2019;81:585–605
- 6. Sheetz M. Mechanobiology in cardiac mechanics Biophys Rev. 2021;13(5):583–585
- 7. Yang H, et al The role of mechanosensitive ion channels in the gastrointestinal tract Front Physiol. 2022;13904203
- 8. Delmas P, Coste B. Mechano-gated ion channels in sensory systems Cell. 2013;155(2):278–284
- 9. Jaalouk DE, Lammerding J. Mechanotransduction gone awry Nat Rev Mol Cell Biol. 2009;10(1):63–73
- 10. Joshi V, et al Mechanotransduction in gastrointestinal smooth muscle cells: role of mechanosensitive ion channels Am J Physiol Gastrointest Liver Physiol. 2021;320(5):G897–G906
- 11. Mazet B. Gastrointestinal motility and its enteric actors in mechanosensitivity: past and present Pflugers Arch. 2015;467(1):191–200
- 12. Alcaino C, et al Mechanosensitive piezo channels in the gastrointestinal tract Curr Top Membr. 2017;79:219–244
- 13. Kraichely RE, Farrugia G. Mechanosensitive ion channels in interstitial cells of Cajal and smooth muscle of the gastrointestinal tract Neurogastroenterol Motil. 2007;19(4):245–252
- 14. Patel B, et al Computational analysis of mechanical stress in colonic diverticulosis Sci Rep. 2020;10(1)6014
- 15. Neshatian L, et al Ranolazine inhibits voltage-gated mechanosensitive sodium channels in human colon circular smooth muscle cells Am J Physiol Gastrointest Liver Physiol. 2015;309(6):G506–G512
- 16. Feng B, Guo T. Visceral pain from colon and rectum: the mechanotransduction and biomechanics J Neural Transm (Vienna). 2020;127(4):415–429
- 17. Balemans D, et al Transient receptor potential ion channel function in sensory transduction and cellular signaling cascades underlying visceral hypersensitivity Am J Physiol Gastrointest Liver Physiol. 2017;312(6):G635–G648
- 18. Page AJ, Li H. Meal-sensing signaling pathways in functional dyspepsia Front Syst Neurosci. 2018;1210
- 19. Fernández-Sánchez ME, et al Mechanical induction of the tumorigenic β-catenin pathway by tumour growth pressure Nature. 2015;523(7558):92–95
- 20. Moriggi M, et al Contribution of extracellular matrix and signal mechanotransduction to epithelial cell damage in inflammatory bowel disease patients: a proteomic study Proteomics. 2017;17(23-24)1700164
- 21. Acosta A, et al Quantitative gastrointestinal and psychological traits associated with obesity and response to weight-loss therapy Gastroenterology. 2015;148(3):537–546
- 22. Brohawn SG, et al Mechanosensitivity is mediated directly by the lipid membrane in TRAAK and TREK1 K+ channels Proc Natl Acad Sci U S A. 2014;111(9):3614–3619
- 23. Friedman SL. Mechanisms of disease: mechanisms of hepatic fibrosis and therapeutic implications Nat Clin Pract Gastroenterol Hepatol. 2004;1(2):98–105
- 24. Olsen AL, et al Hepatic stellate cells require a stiff environment for myofibroblastic differentiation Am J Physiol Gastrointest Liver Physiol. 2011;301(1):G110–G118
- 25. Wells RG. The role of matrix stiffness in regulating cell behavior Hepatology. 2008;47(4):1394–1400
- 26. Romac JM, et al Piezo1 is a mechanically activated ion channel and mediates pressure induced pancreatitis Nat Commun. 2018;9(1)1715
- 27. Yamamoto M, et al A new model of chronic pancreatitis in rats Am J Physiol Gastrointest Liver Physiol. 2006;291(4):G700–G708
- 28. Watanabe S, et al Pressure activates rat pancreatic stellate cells Am J Physiol Gastrointest Liver Physiol. 2004;287(6):G1175–G1181
- 29. Lerch MM, et al Pancreatic duct obstruction triggers acute necrotizing pancreatitis in the opossum Gastroenterology. 1993;104(3):853–861
- 30. Anderson MA, et al Complications of ERCP Gastrointest Endosc. 2012;75(3):467–473
- 31. van Santvoort HC, et al Early endoscopic retrograde cholangiopancreatography in predicted severe acute biliary pancreatitis: a prospective multicenter study Ann Surg. 2009;250(1):68–75
- 32. Gukovskaya AS, et al Mechanisms of cell death after pancreatic duct obstruction in the opossum and the rat Gastroenterology. 1996;110(3):875–884
- 33. Swain SM, et al TRPV4 channel opening mediates pressure-induced pancreatitis initiated by Piezo1 activation J Clin Invest. 2020;130(5):2527–2541
- 34. Swain SM, et al Piezo1-mediated stellate cell activation causes pressure-induced pancreatic fibrosis in mice JCI Insight. 2022;7(8)e158288
- 35. Deivasikamani V, et al Piezo1 channel activation mimics high glucose as a stimulator of insulin release Sci Rep. 2019;9(1)16876
- 36. Ye Y, et al A critical role of the mechanosensor PIEZO1 in glucose-induced insulin secretion in pancreatic β-cells Nat Commun. 2022;13(1)4237
- 37. Coste B, et al Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels Science. 2010;330(6000):55–60
- 38. Wang S, et al Endothelial cation channel PIEZO1 controls blood pressure by mediating flow-induced ATP release J Clin Invest. 2016;126(12):4527–4536
- 39. Swain SM, Liddle RA. Piezo1 acts upstream of TRPV4 to induce pathological changes in endothelial cells due to shear stress J Biol Chem. 2021;296100171
- 40. Syeda R, et al Piezo1 channels are inherently mechanosensitive Cell Rep. 2016;17(7):1739–1746
- 41. Mercado-Perez A, Beyder A. Gut feelings: mechanosensing in the gastrointestinal tract Nat Rev Gastroenterol Hepatol. 2022;19(5):283–296
- 42. Fang XZ, et al Structure, kinetic properties and biological function of mechanosensitive Piezo channels Cell Biosci. 2021;11(1)13
- 43. Wang L, et al Structure and mechanogating of the mammalian tactile channel PIEZO2 Nature. 2019;573(7773):225–229
- 44. Zhao Q, et al Structure and mechanogating mechanism of the Piezo1 channel Nature. 2018;554(7693):487–492
- 45. Murthy SE, et al Piezos thrive under pressure: mechanically activated ion channels in health and disease Nat Rev Mol Cell Biol. 2017;18(12):771–783
- 46. Murthy SE, et al The mechanosensitive ion channel Piezo2 mediates sensitivity to mechanical pain in mice Sci Transl Med. 2018;10(462)eaat9897
- 47. Szczot M, et al PIEZO2 mediates injury-induced tactile pain in mice and humans Sci Transl Med. 2018;10(462)eaat9892
- 48. Vasileva V, Chubinskiy-Nadezhdin V. Regulation of PIEZO1 channels by lipids and the structural components of extracellular matrix/cell cytoskeleton J Cell Physiol. 2023;238(5):918–930
- 49. Poole K, et al Tuning Piezo ion channels to detect molecular-scale movements relevant for fine touch Nat Commun. 2014;53520
- 50. Del Mármol JI, et al Piezo1 forms a slowly-inactivating mechanosensory channel in mouse embryonic stem cells Elife. 2018;7e33149
- 51. Romero LO, et al Dietary fatty acids fine-tune Piezo1 mechanical response Nat Commun. 2019;10(1)1200
- 52. Wang B, et al Mechanosensitive ion channel Piezo1 activated by matrix stiffness regulates oxidative stress-induced senescence and apoptosis in human intervertebral disc degeneration Oxid Med Cell Longev. 2021;20218884922
- 53. Gaub BM, Müller DJ. Mechanical stimulation of Piezo1 receptors depends on extracellular matrix proteins and directionality of force Nano Lett. 2017;17(3):2064–2072
- 54. Lopez-Cavestany M, et al Matrix stiffness induces epithelial-to-mesenchymal transition via Piezo1-regulated calcium flux in prostate cancer cells iScience. 2023;26(4)106275
- 55. Lee W, et al Inflammatory signaling sensitizes Piezo1 mechanotransduction in articular chondrocytes as a pathogenic feed-forward mechanism in osteoarthritis Proc Natl Acad Sci U S A. 2021;118(13)e2001611118
- 56. Zhao X, et al Mechanosensitive Piezo1 channels mediate renal fibrosis JCI Insight. 2022;7(7)e152330
- 57. Dalghi MG, et al Expression and distribution of PIEZO1 in the mouse urinary tract Am J Physiol Renal Physiol. 2019;317(2):F303–F321
- 58. Cahalan SM, et al Piezo1 links mechanical forces to red blood cell volume Elife. 2015;4e07370
- 59. Solis AG, et al Mechanosensation of cyclical force by PIEZO1 is essential for innate immunity Nature. 2019;573(7772):69–74
- 60. Gudipaty SA, et al Mechanical stretch triggers rapid epithelial cell division through Piezo1 Nature. 2017;543(7643):118–121
- 61. Ranade SS, et al Piezo1, a mechanically activated ion channel, is required for vascular development in mice Proc Natl Acad Sci U S A. 2014;111(28):10347–10352
- 62. Li J, et al Piezo1 integration of vascular architecture with physiological force Nature. 2014;515(7526):279–282
- 63. Nonomura K, et al Mechanically activated ion channel PIEZO1 is required for lymphatic valve formation Proc Natl Acad Sci U S A. 2018;115(50):12817–12822
- 64. Lhomme A, et al Stretch-activated Piezo1 channel in endothelial cells relaxes mouse intrapulmonary arteries Am J Respir Cell Mol Biol. 2019;60(6):650–658
- 65. John L, et al The Piezo1 cation channel mediates uterine artery shear stress mechanotransduction and vasodilation during rat pregnancy Am J Physiol Heart Circ Physiol. 2018;315(4):H1019–H1026
- 66. Zeng WZ, et al PIEZOs mediate neuronal sensing of blood pressure and the baroreceptor reflex Science. 2018;362(6413):464–467
- 67. Faucherre A, et al Piezo1 plays a role in erythrocyte volume homeostasis Haematologica. 2014;99(1):70–75
- 68. Pathak MM, et al Stretch-activated ion channel Piezo1 directs lineage choice in human neural stem cells Proc Natl Acad Sci U S A. 2014;111(45):16148–16153
- 69. Velasco-Estevez M, et al Inhibition of Piezo1 attenuates demyelination in the central nervous system Glia. 2020;68(2):356–375
- 70. Wang F, et al Mechanosensitive ion channel Piezo2 is important for enterochromaffin cell response to mechanical forces J Physiol. 2017;595(1):79–91
- 71. Qin L, et al Roles of mechanosensitive channel Piezo1/2 proteins in skeleton and other tissues Bone Res. 2021;9(1)44
- 72. Lee W, et al Synergy between Piezo1 and Piezo2 channels confers high-strain mechanosensitivity to articular cartilage Proc Natl Acad Sci U S A. 2014;111(47):E5114–E5122
- 73. Hasegawa K, et al YAP signaling induces PIEZO1 to promote oral squamous cell carcinoma cell proliferation J Pathol. 2021;253(1):80–93
- 74. Aykut B, et al Targeting Piezo1 unleashes innate immunity against cancer and infectious disease Sci Immunol. 2020;5(50)eabb5168
- 75. Zhang M, et al Mechanically activated piezo channels mediate touch and suppress acute mechanical pain response in mice Cell Rep. 2019;26(6):1419–1431
- 76. Woo SH, et al Piezo2 is required for Merkel-cell mechanotransduction Nature. 2014;509(7502):622–626
- 77. Nonomura K, et al Piezo2 senses airway stretch and mediates lung inflation-induced apnoea Nature. 2017;541(7636):176–181
- 78. Li M, et al Activation of Piezo1 contributes to matrix stiffness-induced angiogenesis in hepatocellular carcinoma Cancer Commun (Lond). 2022;42(11):1162–1184
- 79. Xiong Y, et al Piezo1 activation facilitates ovarian cancer metastasis via Hippo/YAP signaling axis Channels (Austin). 2022;16(1):159–166
- 80. Zhou W, et al Identification of PIEZO1 as a potential prognostic marker in gliomas Sci Rep. 2020;10(1)16121
- 81. Zhu B, et al Piezo 1 activation facilitates cholangiocarcinoma metastasis via Hippo/YAP signaling axis Mol Ther Nucleic Acids. 2021;24:241–252
- 82. Otsuki M, et al Animal models of chronic pancreatitis Gastroenterol Res Pract. 2010;2010403295
- 83. Syeda R, et al Chemical activation of the mechanotransduction channel Piezo1 Elife. 2015;4e07369
- 84. Gerasimenko JV, et al The role of Ca2+ in the pathophysiology of pancreatitis J Physiol. 2014;592(2):269–280
- 85. Raraty M, et al Calcium-dependent enzyme activation and vacuole formation in the apical granular region of pancreatic acinar cells Proc Natl Acad Sci U S A. 2000;97(24):13126–13131
- 86. Bae C, et al The mechanosensitive ion channel Piezo1 is inhibited by the peptide GsMTx4 Biochemistry. 2011;50(29):6295–6300
- 87. Vriens J, et al Cell swelling, heat, and chemical agonists use distinct pathways for the activation of the cation channel TRPV4 Proc Natl Acad Sci U S A. 2004;101(1):396–401
- 88. Watanabe H, et al Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels Nature. 2003;424(6947):434–438
- 89. Steer ML, et al Chronic pancreatitis N Engl J Med. 1995;332(22):1482–1490
- 90. Bradley EL 3rd. Pancreatic duct pressure in chronic pancreatitis Am J Surg. 1982;144(3):313–316
- 91. Gupte A, et al Chronic pancreatitis BMJ. 2018;361k2126
- 92. Schmid-Kotsas A, et al Lipopolysaccharide-activated macrophages stimulate the synthesis of collagen type I and C-fibronectin in cultured pancreatic stellate cells Am J Pathol. 1999;155(5):1749–1758
- 93. Bachem MG, et al Pancreatic carcinoma cells induce fibrosis by stimulating proliferation and matrix synthesis of stellate cells Gastroenterology. 2005;128(4):907–921
- 94. Omary MB, et al The pancreatic stellate cell: a star on the rise in pancreatic diseases J Clin Invest. 2007;117(1):50–59
- 95. Sendler M, et al Complement component 5 mediates development of fibrosis, via activation of stellate cells, in 2 mouse models of chronic pancreatitis Gastroenterology. 2015;149(3):765–776
- 96. Klöppel G, Maillet B. Chronic pancreatitis: evolution of the disease Hepatogastroenterology. 1991;38(5):408–412
- 97. Lew D, et al Chronic pancreatitis: current status and challenges for prevention and treatment Dig Dis Sci. 2017;62(7):1702–1712
- 98. Kalas MA, et al Vascular complications of pancreatitis World J Clin Cases. 2022;10(22):7665–7673
- 99. Chandan S, et al Use of therapeutic anticoagulation in splanchnic vein thrombosis associated with acute pancreatitis: a systematic review and meta-analysis Ann Gastroenterol. 2021;34(6):862–871
- 100. Nadkarni NA, et al Splanchnic venous thrombosis and pancreatitis Pancreas. 2013;42(6):924–931
- 101. Easler J, et al Portosplenomesenteric venous thrombosis in patients with acute pancreatitis is associated with pancreatic necrosis and usually has a benign course Clin Gastroenterol Hepatol. 2014;12(5):854–862
- 102. Wang S, et al Mechanosensation by endothelial PIEZO1 is required for leukocyte diapedesis Blood. 2022;140(3):171–183
- 103. Fuentes IM, Christianson JA. Ion channels, ion channel receptors, and visceral hypersensitivity in irritable bowel syndrome Neurogastroenterol Motil. 2016;28(11):1613–1618
- 104. Smith TK, et al Recent advances in enteric neurobiology: mechanosensitive interneurons Neurogastroenterol Motil. 2007;19(11):869–878
- 105. Beyder A, Farrugia G. Ion channelopathies in functional GI disorders Am J Physiol Gastrointest Liver Physiol. 2016;311(4):G581–G586
- 106. Mazzuoli-Weber G, et al Piezo proteins: incidence and abundance in the enteric nervous system. Is there a link with mechanosensitivity? Cell Tissue Res. 2019;375(3):605–618
- 107. Bartoli F, et al Endothelial Piezo1 sustains muscle capillary density and contributes to physical activity J Clin Invest. 2022;132(5)e141775
- 108. Franco A Jr, Lansman JB. Stretch-sensitive channels in developing muscle cells from a mouse cell line J Physiol. 1990;427:361–380
- 109. Pérez-González C, et al Digesting the mechanobiology of the intestinal epithelium Curr Opin Genet Dev. 2022;72:82–90
- 110. Xu Y, et al Mechanical stimulation activates Piezo1 to promote mucin2 expression in goblet cells J Gastroenterol Hepatol. 2021;36(11):3127–3139
- 111. Alcaino C, et al A population of gut epithelial enterochromaffin cells is mechanosensitive and requires Piezo2 to convert force into serotonin release Proc Natl Acad Sci U S A. 2018;115(32):E7632–E7641
- 112. He L, et al Mechanical regulation of stem-cell differentiation by the stretch-activated Piezo channel Nature. 2018;555(7694):103–106
- 113. Eisenhoffer GT, et al Crowding induces live cell extrusion to maintain homeostatic cell numbers in epithelia Nature. 2012;484(7395):546–549
- 114. Xie Z, et al Piezo2 channels expressed by colon-innervating TRPV1-lineage neurons mediate visceral mechanical hypersensitivity Neuron. 2023;111(4):526–538
- 115. Chesler AT, et al The role of PIEZO2 in human mechanosensation N Engl J Med. 2016;375(14):1355–1364
- 116. Ranade SS, et al Piezo2 is the major transducer of mechanical forces for touch sensation in mice Nature. 2014;516(7529):121–125
- 117. Hill RZ, et al PIEZO1 transduces mechanical itch in mice Nature. 2022;607(7917):104–110
- 118. Wang J, et al PIEZO1 is selectively expressed in small diameter mouse DRG neurons distinct from neurons strongly expressing TRPV1 Front Mol Neurosci. 2019;12178
- 119. Wang P, et al Visceral mechano-sensing neurons control Drosophila feeding by using piezo as a sensor Neuron. 2020;108(4):640–650
- 120. Min S, et al Control of feeding by Piezo-mediated gut mechanosensation in Drosophila Elife. 2021;10e63049
- 121. Oh Y, et al Periphery signals generated by Piezo-mediated stomach stretch and Neuromedin-mediated glucose load regulate the Drosophila brain nutrient sensor Neuron. 2021;109(12):1979–1995
- 122. Goyal RK, Chaudhury A. Physiology of normal esophageal motility J Clin Gastroenterol. 2008;42(5):610–619
- 123. Lowenstein ED, et al Prox2 and Runx3 vagal sensory neurons regulate esophageal motility Neuron. 2023;111(14):2184–2200
- 124. Chin A, et al The role of mechanical forces and adenosine in the regulation of intestinal enterochromaffin cell serotonin secretion Am J Physiol Gastrointest Liver Physiol. 2012;302(3):G397–G405
- 125. Bellono NW, et al Enterochromaffin cells are gut chemosensors that couple to sensory neural pathways Cell. 2017;170(1):185–198
- 126. Crowell MD. Role of serotonin in the pathophysiology of the irritable bowel syndrome Br J Pharmacol. 2004;141(8):1285–1293
- 127. Houghton LA, et al Alosetron, a 5-HT3 receptor antagonist, delays colonic transit in patients with irritable bowel syndrome and healthy volunteers Aliment Pharmacol Ther. 2000;14(6):775–782
- 128. Bouras EP, et al Selective stimulation of colonic transit by the benzofuran 5HT4 agonist, prucalopride, in healthy humans Gut. 1999;44(5):682–686
- 129. Shin SM, et al Piezo2 mechanosensitive ion channel is located to sensory neurons and nonneuronal cells in rat peripheral sensory pathway: implications in pain Pain. 2021;162(11):2750–2768
- 130. Treichel AJ, et al Specialized mechanosensory epithelial cells in mouse gut intrinsic tactile sensitivity Gastroenterology. 2022;162(2):535–547.e13
- 131. Yang J, et al The potential role of Piezo2 in the mediation of visceral sensation Neurosci Lett. 2016;630:158–163
- 132. Ritchie J. Pain from distension of the pelvic colon by inflating a balloon in the irritable colon syndrome Gut. 1973;14(2):125–132
- 133. Dinis P, et al Anandamide-evoked activation of vanilloid receptor 1 contributes to the development of bladder hyperreflexia and nociceptive transmission to spinal dorsal horn neurons in cystitis J Neurosci. 2004;24(50):11253–11263
- 134. Jones RC, et al The mechanosensitivity of mouse colon afferent fibers and their sensitization by inflammatory mediators require transient receptor potential vanilloid 1 and acid-sensing ion channel 3 J Neurosci. 2005;25(47):10981–10989
- 135. Schwartz ES, et al TRPV1 and TRPA1 antagonists prevent the transition of acute to chronic inflammation and pain in chronic pancreatitis J Neurosci. 2013;33(13):5603–5611
- 136. Holzer P. TRPV1: a new target for treatment of visceral pain in IBS? Gut. 2008;57(7):882–884
- 137. Yang XN, et al Piezo1 is as a novel trefoil factor family 1 binding protein that promotes gastric cancer cell mobility in vitro Dig Dis Sci. 2014;59(7):1428–1435
- 138. Ning W, et al Differentiated daughter cells regulate stem cell proliferation and fate through intra-tissue tension Cell Stem Cell. 2021;28(3):436–452
- 139. Gao L, et al Suppression of esophageal squamous cell carcinoma development by mechanosensitive protein Piezo1 downregulation ACS Omega. 2021;6(15):10196–10206
- 140. Qin X, et al High-throughput membrane-anchored proteome screening reveals PIEZO1 as a promising antibody-drug target for human esophageal squamous cell carcinoma Cancer Med. 2022;11(19):3700–3713
- 141. Perez-Gonzalez NA, et al YAP and TAZ regulate cell volume J Cell Biol. 2019;218(10):3472–3488
- 142. Castellan M, et al Single-cell analyses reveal YAP/TAZ as regulators of stemness and cell plasticity in glioblastoma Nat Cancer. 2021;2(2):174–188
- 143. Sun Y, et al The function of Piezo1 in colon cancer metastasis and its potential regulatory mechanism J Cancer Res Clin Oncol. 2020;146(5):1139–1152
- 144. Ioannou M, et al HIF-1α in colorectal carcinoma: review of the literature J BUON. 2015;20(3):680–689
- 145. Ren T, et al MCU-dependent mitochondrial Ca2+ inhibits NAD+/SIRT3/SOD2 pathway to promote ROS production and metastasis of HCC cells Oncogene. 2017;36(42):5897–5909
- 146. Hope JM, et al Activation of Piezo1 sensitizes cells to TRAIL-mediated apoptosis through mitochondrial outer membrane permeability Cell Death Dis. 2019;10(11)837
- 147. Zhang J, et al PIEZO1 functions as a potential oncogene by promoting cell proliferation and migration in gastric carcinogenesis Mol Carcinog. 2018;57(9):1144–1155
- 148. Li YM, et al Piezo1 promoted hepatocellular carcinoma progression and EMT through activating TGF-β signaling by recruiting Rab5c Cancer Cell Int. 2022;22(1)162
- 149. Huang Z, et al Loss of stretch-activated channels, PIEZOs, accelerates non-small cell lung cancer progression and cell migration Biosci Rep. 2019;39(3)BSR20181679
- 150. Wang Y, et al Immunoregulatory role of the mechanosensitive ion channel Piezo1 in inflammation and cancer Molecules. 2022;28(1)213
- 151. Chakraborty M, et al Mechanical stiffness controls dendritic cell metabolism and function Cell Rep. 2021;34(2)108609
- 152. Wang Y, et al Dendritic cell Piezo1 directs the differentiation of TH1 and Treg cells in cancer Elife. 2022;11e79957
- 153. Yang J, et al Roles of plasmacytoid dendritic cells in gastric cancer Front Oncol. 2022;12818314
- 154. Rihawi K, et al Tumor-associated macrophages and inflammatory microenvironment in gastric cancer: novel translational implications Int J Mol Sci. 2021;22(8)3805
- 155. Gambardella V, et al The role of tumor-associated macrophages in gastric cancer development and their potential as a therapeutic target Cancer Treat Rev. 2020;86102015
- 156. Wang H, et al Tumor-associated macrophages (TAMs) in colorectal cancer (CRC): from mechanism to therapy and prognosis Int J Mol Sci. 2021;22(16)8470
- 157. Liu Q, et al The mechanosensitive ion channel PIEZO1 in intestinal epithelial cells mediates inflammation through the NOD-like receptor 3 pathway in Crohn’s disease Inflamm Bowel Dis. 2023;29(1):103–115
- 158. Choi SH, et al Mechanosensory channel Piezo1 is essential in pathogenic T cell-mediated intestinal inflammation [abstract] J Immunol. 2022;208(1 suppl)113.16
- 159. Liu Y, et al Reprogrammed fecal and mucosa-associated intestinal microbiota and weakened mucus layer in intestinal goblet cell- specific Piezo1-deficient mice Front Cell Infect Microbiol. 2022;121035386
- 160. Konturek PC, et al Stress and the gut: pathophysiology, clinical consequences, diagnostic approach and treatment options J Physiol Pharmacol. 2011;62(6):591–599
- 161. Ilchmann-Diounou H, Menard S. Psychological stress, intestinal barrier dysfunctions, and autoimmune disorders: an overview Front Immunol. 2020;111823
- 162. Rengarajan S, et al A potential role for stress-induced microbial alterations in IgA-associated irritable bowel syndrome with diarrhea Cell Rep Med. 2020;1(7)100124
- 163. Bradesi S, et al Repeated exposure to water avoidance stress in rats: a new model for sustained visceral hyperalgesia Am J Physiol Gastrointest Liver Physiol. 2005;289(1):G42–G53
- 164. Jiang Y, et al Piezo1 regulates intestinal epithelial function by affecting the tight junction protein claudin-1 via the ROCK pathway Life Sci. 2021;275119254
Conflict of interest: The authors have declared that no conflict of interest exists.
Copyright: © 2023, Swain et al. This is an open access article published under the terms of the Creative Commons Attribution 4.0 International License.
Reference information: J Clin Invest. 2023;133(19):e171955. https://doi.org/10.1172/JCI171955.