Introduction
Nitrogen (N) availability is a major factor determining plant growth and productivity. Plants can acquire N through their roots from the soil under inorganic (nitrate and ammonium) and organic (e.g. urea, amino acids, peptides) forms. Although organic forms contribute to plant N nutrition in specific habitats such as in boreal ecosystems (, ), nitrate and ammonium are the universal forms in most soils. In natural soils, their availability is generally low but can also be highly variable depending on various factors including soil physical properties, leaching and microbial activity, which often result in the formation of N depletion areas in the soil (, ). To face such low N availability conditions, plants display elaborate responses to enhance N use efficiency (, , ).
N use efficiency has been defined in multiple ways; however, in general, it can be divided into two components, N utilization efficiency and N acquisition (uptake) efficiency. Uptake activity and root architecture are the major determinants of the acquisition efficiency (reviewed in , , ). The former is facilitated by influx transporters located on the plasma membrane, and the latter by alterations in growth and development in response to local and systemic N signals (, ). Needless to say these two factors are co-ordinated to optimize acquisition. This review aims to summarize recent advances in our understanding of the mechanisms that plants employ to increase N acquisition efficiency under low N availability, with special reference to the regulation of uptake and root architecture in Arabidopsis thaliana (Arabidopsis). In this review, we mostly focus on ‘N limitation (sudden complete deprivation, low or growth-limiting concentrations)’ and ‘heterogeneous supply (nutrient patches simulated by split-root)’.
Regulation of Uptake
Ammonium and nitrate are taken up actively into root cells by different sets of plasma membrane-localized transporters. Ammonium transport is mediated by transporters of the AMT/MEP/Rh (AMT) superfamily (). Six AMT genes exist in Arabidopsis, all of which encode high-affinity ammonium transporters (, ). For nitrate uptake, two families of transporters, NPF, for NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family (previously named the NRT1/PTR family) and NRT2, have been identified (, ). In Arabidopsis, there are 53 and seven members in the NPF and NRT2 families, respectively. The NPF members studied so far have a low affinity for nitrate (), except AtNPF6.3 (NRT1.1), which has dual-affinity transport and nitrate-sensing functions (). NRT2 members are high-affinity nitrate transporters, and most of them require another component, NAR2 (NRT3), to mediate nitrate transport (, , ). A series of genetic and physiological studies have shown that high-affinity transporters plays a central part in efficient N uptake under low availability (, ).
Molecular Basis of Ammonium Uptake
Five of the six AMT genes in Arabidopsis, namely AtAMT1;1, AtAMT1;2, AtAMT1;3, AtAMT1;5 and AtAMT2;1, are expressed in roots, and their transcript levels are up-regulated under N limitation (). Ammonium influx studies using triple and quadruple mutants showed that AtAMT1;1, AtAMT1;2 and AtAMT1;3 are additively responsible for about 90% of high-affinity uptake capacity under N limitation, and that AtAMT1;5 most probably accounts for the remaining capacity (, ). AtAMT1;1, AtAMT1;3 and AtAMT1;5 are expressed chiefly at the root tip and in epidermal cells, while AtAMT1;2 is localized in the endodermis and cortex. All AtAMTs have different ammonium affinity levels and transport capacities indicative of their roles in planta. AtAMT1;1, AtAMT1;3 and AtAMT1;5 act to absorb ammonium directly from the soil, and AtAMT1;2 transports apoplastic ammonium into the cell (, ). These lines of evidence illustrate how important it is for the regulation of multiple transporters with appropriate substrate affinity and capacity to be co-ordinated for effective ammonium uptake under low availability.
Although AMT expression is derepressed by N limitation in Arabidopsis, ammonium-inducible expression has been reported in other plants such as rice, tomato and maize (, ). Furthermore, the rice and poplar genomes contain 10 and 14 putative AMT genes, respectively (, ), suggesting that the co-ordinated regulation patterns of AMT genes could be quite intricate depending on plant species and habitat.
Molecular Basis of Nitrate Uptake
Among seven NRT2 genes in Arabidopsis, AtNRT2.1, AtNRT2.2, AtNRT2.4 and AtNRT2.5 are expressed in the roots of N-deprived plants. Analysis of a quadruple mutant revealed that these four NRT2 transporters account for approximately 95% of high-affinity nitrate influx activity under N limitation, AtNRT2.1 being the major contributor (). Recent studies suggest that the spatio-temporal distribution of these four AtNRT2 transporters is critical for efficient nitrate uptake to sustain growth under low N availability (Fig. 1; , ). During N deprivation, the expression of AtNRT2.1 is transiently derepressed in the cortex cells of older parts of primary and lateral roots (). In contrast, the transcript levels of AtNRT2.4 and AtNRT2.5 increase during N deprivation over time in the epidermal cells of young primary and lateral roots (, , ). These spatial expression patterns indicate that AtNRT2.4 and AtNRT2.5 are responsible for nitrate uptake from the soil, and AtNRT2.1 plays a role in apoplastic nitrate absorption. Although AtNRT2.4 and AtNRT2.5 are expressed in the same cell types, the former is predominant in young seedlings and the latter in adult plants (Fig. 1; , ). In addition, AtNRT2.4 was suggested to have much higher affinity for nitrate than AtNRT2.1 (). Although a dual-affinity transporter AtNPF6.3 is also expressed in roots under N limitation, its direct contribution to high-affinity nitrate transport under N limitation seems to be minor, maybe even non-existent (). The existence of a high-affinity efflux transport system for xylem loading of nitrate (Fig. 1A) has been suggested from the phenotype of atnrt1.5, which is a mutant of a low-affinity efflux transporter responsible for xylem loading of nitrate (). However, the transporter gene(s) involved in the system is(are) still unknown.
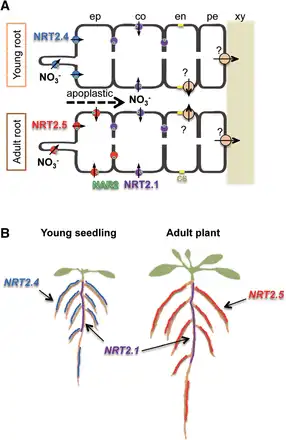
Fig. 1
Schematic illustration summarizing the function of NRT2 transporters in Arabidopsis roots under low N availability. Spatial and temporal localization of AtNRT2.1 (NRT2.1, purple), AtNRT2.4 (NRT2.4, blue) and AtNRT2.5 (NRT2.5, red) in (A) root tissues and (B) whole root systems under low N availability. (A) The NRT2.4/NAR2 complex is localized to the outer (soil) side of the epidermal cells of the roots of young seedlings. The NRT2.5/NAR2 complex is expressed in the epidermal cells of the roots of adult plants. NRT2.4 and NRT2.5 are responsible for nitrate uptake directly from the soil. Nitrate can apoplastically penetrate toward cortex cells to be absorbed by the NRT2.1/NAR2 complex. NAR2 (AtNAR2.1) is shown as green circles. Orange circles indicate a putative high-affinity exporter involved in xylem loading of nitrate. (B) NRT2.1 is strongly expressed in the older part of the root system, while NRT2.4 and NRT2.5 are preferentially expressed in the younger part of the roots of young seedlings and adult plants, respectively. ep, epidermis; co, cortex; en, endodermis; pe, pericycle; xy, xylem; cs, casparian strip
The NRT2 family genes have also been investigated in other plant species, and many N-limitation-inducible genes have been identified (, ). Biochemically, they seem to act as high-affinity nitrate transporters (, ). However, functional characterization of these genes in planta remains to be carried out.
Regulation of High-Affinity N Transporter Genes at the Transcript Level
Generally the expression of genes encoding high-affinity transporters of mineral nutrients is induced (or derepressed) under low substrate availability. This is also the case for all AtAMT1 and AtNRT2 genes involved in uptake, as described above. A reduction of the internal pool of glutamine and/or derived metabolites is thought to be one of the signals for induction (, , , ). Recently AtNPF6.3 was shown to be involved in the negative regulation of induction under high N availability. In the knockout mutant of AtNPF6.3 (chl1-5) grown under high N availability, the expression of high-affinity N transporter genes including AtNRT2.1, AtNRT2.4 and AtAMT1;3 was derepressed (Fig. 2A; , ). Phosphorylation of the threonine (T) 101 residue (T101) plays a role in this regulation. Introducing the phosphomimetic mutant form AtNPF6.3T101D into chl1-5 restored repression, but the non-phosphorylatable mutant form AtNPF6.3T101A did not. However, how low N availability is sensed and how the signal is transduced through phosphorylation of AtNPF6.3 is still unknown.
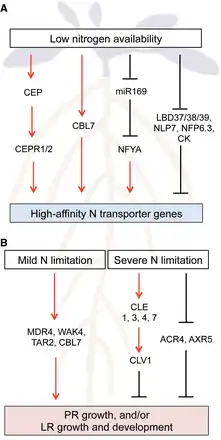
Fig. 2
A model of low N availability signaling pathways involved in the regulation of high-affinity N transporter gene expression and root architecture in Arabidopsis. Signaling pathways regulating (A) the expression of high-affinity N transporter genes (AtNRT2 and AtAMT genes) and (B) primary root (PR) growth, and/or lateral root (LR) growth and development under low N availability are depicted. Only pathways described in this review are shown. Red arrows and black blunted lines indicate positive and negative interactions, respectively. CK, cytokinin
LATERAL ORGAN BOUNDARY DOMAIN (LBD) family transcription factors (LBD37/38/39) and members of the RWP-RK family transcription factor NIN-LIKE PROTEIN (NLP) are implicated in the regulation of high-affinity N transporter genes under low N availability in Arabidopsis. Overexpression of LBD37/38/39 suppressed some N limitation responses, including the induction of AtNRT2.1, AtNRT2.2 and AtNRT2.5 (Fig. 2A). The expression of LBD37/38/39 is low under N limitation, while it is high under N sufficiency, indicating that LBD37/38/39 function as repressors of N limitation responses under N sufficiency (). Recently NLP6 and NLP7 were shown to act as master regulators of the primary nitrate response in Arabidopsis (, , ). Interestingly, nlp7 mutants display constitutive N limitation responses, including induction of AtAMT1;5 and AtNRT2.5 (), indicating that NLP7 also plays a role in the repression of N limitation responses. Indeed, chromatin immunoprecipitation analysis revealed that NLP7 is bound to high-affinity N transporter genes such as AtAMT1;1, AtNRT2.1 and AtNRT2.5, showing direct regulation (Fig. 2A; ).
The expression of high-affinity N transporter genes is also regulated by systemic N signals. Cytokinins, a class of plant hormones, have been proposed to act as both local and systemic signals co-ordinating N demand and acquisition (). A positive correlation between tissue and vascular cytokinin contents and N availability has been reported in many plant species (, , ). Exogenous application of cytokinins represses AtNRT2 genes in N-deprived roots (), suggesting that cytokinins act as N sufficiency signals to suppress nitrate uptake by the roots (Fig. 2A). Consistently, split-root experiments showed that cytokinins are involved in systemic N deficiency (demand) signaling to regulate AtNRT2.1, AtNAR2 and AtNRT2.4 under heterologous N supply ().
MicroRNAs (miRNAs) are also implicated in local and systemic N signaling. Although a number of miRNAs responsive to low N availability have been identified in various plant species (, , ), few of them have been characterized. Arabidopsis miR169, which targets NUCLEAR FACTOR Y, SUBUNIT A (NFYA) family members, is so far the only one shown be involved in the regulation of N uptake under low N availability. miR169 is down-regulated by N limitation, while the transcript levels of NFYA genes are up-regulated (, ). Transgenic plants overexpressing MIR169a display reduced levels of NFYA transcripts and show hypersensitivity to N limitation, which is associated with decreased AtNRT2.1 expression (). The promoter region of AtNRT2.1 contains an NFY-binding consensus sequence, suggesting that miR169 regulates AtNRT2.1 through NFY (Fig. 2A). Interestingly, miR169 was abundantly detected in rapeseed phloem sap (), implying its role as a systemic N signal. Recently identified C-TERMINALLY ENCODED PEPTIDEs (CEPs) as systemic N deficiency (demand) signals under heterologous N supply in Arabidopsis. CEPs are small peptides produced in the N-deprived side of the root. After translocation to the shoot, CEPs are perceived by two leucine-rich repeat receptor kinases CEP RECEPTOR1/2 (CEPR1/2) that increase AtNRT2.1 and AtNPF6.3 expression and nitrate uptake in the other side of the root (Fig. 2A; ). However, the mechanism whereby shoot-located CEPR1/2 activates AtNRT2.1 and AtNPF6.3 expression in the root remains to be elucidated.
Stability of the mRNA encoding high-affinity N transporter genes can also regulate efficient uptake. Transcript stability of AtAMT1;1 is regulated by N availability (). The AtAMT1;1 transcript level driven by the Cauliflower mosaic virus 35S RNA promoter accumulated under N limitation in tobacco, while its level decreased after resupply of ammonium or nitrate.
Regulation of High-Affinity N Transporters at Post-Translational Levels
Post-translational regulation of transporters would be of benefit because it enables plants to respond immediately to sudden changes in N availability. The best characterized transporters for post-translational regulation include AtAMT1;1 and AtAMT1;3. The transport activity of AtAMT1;1 and AtAMT1;3 is regulated by the phosphorylation status of a threonine residue in the cytosolic C-terminal domain. Phosphorylation, which represses transport activity, occurs when plants are grown with ammonium, but does not occur during N limitation (). AtNPF6.3 is also regulated by phosphorylation in response to nitrate availability. AtNPF6.3 is phosphorylated at T101 by CBL-INTERACTING PROTEIN KINASE 23 (CIPK23) under low nitrate to function as a high-affinity transporter, while it is dephosphorylated under high nitrate to be a low-affinity transporter (, ). Crystallography suggested that dimer decoupling caused by T101 phosphorylation is relevant to the change in affinity for nitrate (, ).
Several studies suggest that AtNRT2s are also regulated post-translationally. When AtNRT2.1 was expressed under the control of the 35S promoter, the activity of high-affinity transporters was repressed by high N supply, even though mRNAs accumulated constantly. Under the same conditions, AtNRT2.1 and AtNAR2.1 protein levels were not correlated with the activity (, ), implying post-translational regulation. Regulation could involve dissociation of the AtNRT2.1/AtNAR2.1 complex (), cleavage of the C-terminus () and phosphorylation of AtNRT2.1 (), though the relevance of such regulation to nitrate uptake under low N availability remains to be demonstrated.
The polar localization of transporters is believed to be important for efficient nutrient uptake. Transporters of various micronutrients, such as the boron transporters AtBOR1 and AtNIP5;1, the silicon transporters OsLsi1 and OsLsi2, a manganese transporter OsNRAMP5, and an iron transporter AtIRT1 display lateral polarity (, , ). However, reports of polar localization of macronutrient transporters are scarce. Among N transporters, AtNRT2.4 is so far the only one shown to have a polar localization. Its localization to the outer (soil) side of epidermal cells is likely to be important for AtNRT2.4 to operate with much higher affinity than AtNRT2.1 (Fig. 1A; ). Although the molecular mechanism of AtNRT2.4 polar localization is unknown, endocytic trafficking between the plasma membrane and endosomes may be a common underlying mechanism for the establishment of polarity for mineral transporters (, ). Consistently, single-particle fluorescence imaging revealed that AtAMT1;3 on the plasma membrane is regulated by clustering and endocytosis under high ammonium ().
Regulation of Root Architecture Under Low N Availability
Plants can improve nutrient uptake by modulating root growth and architecture. By increasing the total absorptive surface of the root system and directing growth toward nutrient-rich patches of the soil, plants are able to adapt to nutrient availability in the soil. Taking into account the high mobility of nitrate in the soil and the restricted amount of nitrate available within a given soil area, the definition of the most efficient root architecture might not be that obvious and may vary depending on plant species, soil type and other environmental parameters (). Modeling approaches suggest that efficient nitrate capture results from a trade-off between the speed of N acquisition and the total volume of soil explored ().
Several different aspects of N-dependent modulation of root morphology are reported and discussed in recent reviews (, ). The nutritional status of the plant as well as local signals detected by the roots trigger morphological changes in the overall root system. Nitrate, ammonium and glutamate act locally on the roots and induce different morphological responses (, ). The modification of root growth under N limitation depends on the strength of N limitation and on other environmental conditions, such as light intensity and day length. Primary and lateral root length is increased under mild N limitation (, ), whereas total root development is delayed under severe N limitation, leading to short primary roots and a proportionally reduced number of lateral roots ().
A survey of the expression of about 100 root development-related genes in the publicly available transcriptome data of Arabidopsis () revealed that N limitation induces the expression of the WALL ASSOCIATED KINASE 4 (WAK4) and of the shootward auxin transporter MULTIDRUG RESISTANCE 4/P-GLYCOPROTEIN 4 (MDR4/PGP4). Both genes stimulate primary and lateral root growth (, ) and might be involved in the response to mild N limitation (Fig. 2B). On the other hand, down-regulation of the expression of ARABIDOPSIS CRINKLY 4 (ACR4) and AUXIN RESISTANT 5 (AXR5), both involved in lateral root formation (, ), might indicate that these genes are involved in the reduction of lateral root formation under severe N limitation (Fig. 2B).
The regulatory mechanisms involved in the responses to changing N availability are multiple, and suggest a complex regulatory network also involving hormonal regulation. In particular, auxin plays a determining role not only in the local nitrate response (), but also in the response to N limitation in Arabidopsis (). Under mild N limitation, auxin accumulates in the non-emerged lateral root primordia with more than three cell layers, leading to increased lateral root growth. This is accompanied by an increase in the expression of the auxin biosynthesis gene TRYPTOPHAN AMINOTRANSFERASE RELATED 2 (TAR2) in the pericycle and the vasculature of the mature root zone near the root tip. Loss of function of TAR2 impairs auxin accumulation and lateral root growth. Conversely, overexpression of TAR2 increases lateral root numbers under both high and low N supply. This causal correlation between TAR2 and auxin accumulation and lateral root number is thus part of the regulation of root architecture under low N availability (Fig. 2B).
Also based on the up-regulation of gene expression under N limitation, members of the CLAVATA3/ESR-RELATED (CLE) gene family are part of a (probably) different regulatory mechanism that restrictively controls the expansion of the lateral root system in N-limited environments in Arabidopsis (). Expression of peptides CLE1, 3, 4 and 7 is induced under N limitation chiefly in the root pericycle cells. Their overexpression leads to reduced lateral root growth. CLE peptides are ligands of the CLAVATA1 (CLV1) leucine-rich repeat receptor-like kinases. CLV1, the receptor of CLE3, is expressed in phloem companion cells. The overexpression of CLE3 in a clv1 mutant background did not modify root architecture. In addition, the clv1 mutant was impaired for N-regulated lateral root primordia outgrowth. This signaling module is localized in the root vasculature, and acts under severe N limitation as a mechanism to prevent the expansion of the lateral root system into N-poor environments (Fig. 2B).
A link with calcium signaling exists, as the CALCINEURIN B-LIKE PROTEIN 7 (CBL7) is involved in the regulation of root growth upon nitrate limitation in Arabidopsis (). cbl7 mutants display reduced primary root growth specifically under very low N availability (Fig. 2B). This growth phenotype is accompanied by a decreased root nitrate content and reduced expression of AtNRT2.4 and AtNRT2.5. The corresponding regulatory mechanism still needs to be further revealed. However, calcium (Ca2+) has recently been shown to act as a secondary messenger in the primary nitrate response in Arabidopsis ().
Perspectives
Because of the environmental and economic impacts of excessive fertilizer use, there is a growing demand for new crop varieties suited for low-input sustainable agriculture (). Considering the importance of N, improving N acquisition efficiency, whatever the breeding technique may be, could be one promising approach to generate such crops. Thus it is imperative to identify the mechanisms that plants inherently possess to enhance N acquisition under low N availability. As summarized in this review, there has been a great deal of progress in our understanding of the genes and signaling pathways that regulate N uptake and root architecture (Figs. 1, 2). Although high-affinity N transporter genes have been thoroughly characterized in Arabidopsis, it is still unclear how N acquisition could be improved by manipulating high-affinity N transporters. Several attempts have been made to improve N acquisition efficiency by constitutively expressing a high-affinity transporter gene, without much success (, , , ). Since it has become evident that spatio-temporal orchestration of multiple transporters is a key mechanism underlying efficient uptake of N (Fig. 1) and of other nutrients (, ), co-ordinated manipulation of multiple transporters may be an effective strategy to improve N acquisition. Thus, it will be of great interest to investigate the mechanisms whereby this orchestration is achieved.
Positive coincidences between quantitative trait loci (QTLs) for N uptake and root architecture have been observed in maize (, ), suggesting that optimal root architecture would improve N acquisition. However, our understanding of molecular components that regulate the root architecture in response to N availability is mostly limited to Arabidopsis. Whether the function of the components is equivalent in other plants, especially in monocots, needs to be tested. Furthermore, one future challenge will be to understand how the transporter orchestration and the modulation of root architecture are co-ordinated to maximize acquisition efficiency in a fluctuating environment.
Funding
This work was supported by the Japan Society for the Promotion of Science [a Grant-in-Aid for Young Scientists (A) (No. 26712009 to T.K.)]; the National Agency for research (ANR) [grant ANR-14-CE19-0008 to A.K.]; LabEx Saclay Plant Sciences-SPS [for work in the laboratory of A.K. (ANR-10-LABX-0040-SPS)].
Disclosures
The authors have no conflicts of interest to declare.
AMT: ammonium transporter
CBL: calcineurin B-like protein
NRT: nitrate transporter
References
- Araya T., Kubo T., von Wiren N., Takahashi H. (2015) Statistical modeling of nitrogen-dependent modulation of root system architecture in Arabidopsis thaliana. J. Integr. Plant Biol. 58: 254–265.
- Araya T., Miyamoto M., Wibowo J., Suzuki A., Kojima S., Tsuchiya Y.N., et al.. (2014) CLE–CLAVATA1 peptide–receptor signaling module regulates the expansion of plant root systems in a nitrogen-dependent manner. Proc. Natl. Acad. Sci. USA 111: 2029–2034.
- Bao A., Liang Z., Zhao Z., Cai H. (2015) Overexpressing of OsAMT1–3, a high affinity ammonium transporter gene, modifies rice growth and carbon–nitrogen metabolic status. Int. J. Mol. Sci. 16: 9037–9063.
- Barberon M., Dubeaux G., Kolb C., Isono E., Zelazny E., Vert G. (2014) Polarization of IRON-REGULATED TRANSPORTER 1 (IRT1) to the plant–soil interface plays crucial role in metal homeostasis. Proc. Natl. Acad. Sci. USA 111: 8293–8298.
- Bouguyon E., Brun F., Meynard D., Kubeš M., Pervent M., Leran S., et al.. (2015) Multiple mechanisms of nitrate sensing by Arabidopsis nitrate transceptor NRT1.1. Nat. Plant 1: 15015.
- Castaings L., Camargo A., Pocholle D., Gaudon V., Texier Y., Boutet-Mercey S., et al.. (2009) The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. 57: 426–435.
- Coque M., Martin A., Veyrieras J.B., Hirel B., Gallais A. (2008) Genetic variation for N-remobilization and postsilking N-uptake in a set of maize recombinant inbred lines. 3. QTL detection and coincidences. Theor. Appl. Genet. 117: 729–747.
- Couturier J., Montanini B., Martin F., Brun A., Blaudez D., Chalot M. (2007) The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 174: 137–150.
- De Smet I., Vassileva V., De Rybel B., Levesque M.P., Grunewald W., Van Damme D., et al.. (2008) Receptor-like kinase ACR4 restricts formative cell divisions in the Arabidopsis root. Science 322: 594–597.
- Dunbabin V., Diggle A., Rengel Z. (2003) Is there an optimal root architecture for nitrate capture in leaching environments? Plant Cell Environ. 26: 835–844.
- Engelsberger W.R., Schulze W.X. (2012) Nitrate and ammonium lead to distinct global dynamic phosphorylation patterns when resupplied to nitrogen-starved Arabidopsis seedlings. Plant J. 69: 978–995.
- Forde B.G. (2014) Nitrogen signalling pathways shaping root system architecture: an update. Curr. Opin. Plant Biol. 21: 30–36.
- Fraisier V., Gojon A., Tillard P., Daniel-Vedele F. (2000) Constitutive expression of a putative high-affinity nitrate transporter in Nicotiana plumbaginifolia: evidence for post-transcriptional regulation by a reduced nitrogen source. Plant J. 23: 489–496.
- Garnett T., Conn V., Kaiser B.N. (2009) Root based approaches to improving nitrogen use efficiency in plants. Plant Cell Environ. 32: 1272–1283.
- Giehl R.F., Gruber B.D., von Wiren N. (2014) It’s time to make changes: modulation of root system architecture by nutrient signals. J. Exp. Bot. 65: 769–778.
- Glass A.D.M. (2003) Nitrogen use efficiency of crop plants: physiological constraints upon nitrogen absorption. Crit. Rev.Plant Sci. 22: 453–470.
- Glass A.D.M., Kotur Z. (2013) A reevaluation of the role of Arabidopsis NRT1.1 in high-affinity nitrate transport. Plant Physiol. 163: 1103–1106.
- Good A.G., Shrawat A.K., Muench D.G. (2004) Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 9: 597–605.
- Gruber B.D., Giehl R.F.H., Friedel S., von Wiren N. (2013) Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 163: 161–179.
- Gu C.S., Zhang X.X., Jiang J.F., Guan Z.Y., Zhao S., Fang W.M., et al.. (2014) Chrysanthemum CmNAR2 interacts with CmNRT2 in the control of nitrate uptake. Sci. Rep. 4: 5833.
- Gu R., Duan F., An X., Zhang F., von Wiren N., Yuan L. (2013) Characterization of AMT-mediated high-affinity ammonium uptake in roots of maize (Zea mays L.). Plant Cell Physiol. 54: 1515–1524.
- Guo T.C., Xuan H.M., Yang Y.Y., Wang L.N., Wei L.T., Wang Y.H., et al.. (2014) Transcription analysis of genes encoding the wheat root transporter NRT1 and NRT2 families during nitrogen starvation. J. Plant Growth Regul. 33: 837–848.
- Hermans C., Hammond J.P., White P.J., Verbruggen N. (2006) How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 11: 610–617.
- Hirose N., Takei K., Kuroha T., Kamada-Nobusada T., Hayashi H., Sakakibara H. (2008) Regulation of cytokinin biosynthesis, compartmentalization and translocation. J. Exp. Bot. 59: 75–83.
- Ho C.H., Lin S.H., Hu H.C., Tsay Y.F. (2009) CHL1 functions as a nitrate sensor in plants. Cell 138: 1184–1194.
- Jackson R.B., Caldwell M.M. (1993) The scale of nutrient heterogeneity around individual plants and its quantification with geostatistics. Ecology 74: 612–614.
- Jones D.L., Kielland K. (2012) Amino acid, peptide and protein mineralization dynamics in a taiga forest soil. Soil Biol. Biochem. 55: 60–69.
- Kamada-Nobusada T., Makita N., Kojima M., Sakakibara H. (2013) Nitrogen-dependent regulation of de novo cytokinin biosynthesis in rice: the role of glutamine metabolism as an additional signal. Plant Cell Physiol. 54: 1881–1893.
- Katayama H., Mori M., Kawamura Y., Tanaka T., Mori M., Hasegawa H. (2009) Production and characterization of transgenic rice plants carrying a high-affinity nitrate transporter gene (OsNRT2.1). Breed. Sci. 59: 237–243.
- Kiba T., Feria-Bourrellier A.B., Lafouge F., Lezhneva L., Boutet-Mercey S., Orsel M., et al.. (2012) The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 24: 245–258.
- Kiba T., Kudo T., Kojima M., Sakakibara H. (2011) Hormonal control of nitrogen acquisition: roles of auxin, abscisic acid, and cytokinin. J. Exp. Bot. 62: 1399–1409.
- Konishi M., Yanagisawa S. (2013) Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 4: 1617.
- Kotur Z., Glass A.D.M. (2015) A 150 kDa plasma membrane complex of AtNRT2.5 and AtNAR2.1 is the major contributor to constitutive high-affinity nitrate influx in Arabidopsis thaliana. Plant Cell Environ. 38: 1490–1502.
- Kotur Z., Mackenzie N., Ramesh S., Tyerman S.D., Kaiser B.N., Glass A.D. (2012) Nitrate transport capacity of the Arabidopsis thaliana NRT2 family members and their interactions with AtNAR2.1. New Phytol. 194: 724–731.
- Krapp A., Berthome R., Orsel M., Mercey-Boutet S., Yu A., Castaings L., et al.. (2011) Arabidopsis roots and shoots show distinct temporal adaptation patterns toward nitrogen starvation. Plant Physiol. 157: 1255–1282.
- Krapp A., David L.C., Chardin C., Girin T., Marmagne A., Leprince A.S., et al.. (2014) Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 65: 789–798.
- Krouk G., Lacombe B., Bielach A., Perrine-Walker F., Malinska K., Mounier E., et al.. (2010) Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 18: 927–937.
- Kumar A., Kaiser B.N., Siddiqi M.Y., Glass A.D.M. (2006) Functional characterisation of OsAMT1.1 overexpression lines of rice, Oryza sativa. Funct. Plant Biol. 33: 339–346.
- Lally D., Ingmire P., Tong H.Y., He Z.H. (2001) Antisense expression of a cell wall-associated protein kinase, WAK4, inhibits cell elongation and alters morphology. Plant Cell 13: 1317–1331.
- Laugier E., Bouguyon E., Mauries A., Tillard P., Gojon A., Lejay L. (2012) Regulation of high-affinity nitrate uptake in roots of Arabidopsis depends predominantly on posttranscriptional control of the NRT2.1/NAR2.1 transport system. Plant Physiol. 158: 1067–1078.
- Lejay L., Tillard P., Lepetit M., Olive F.D., Filleur S., Daniel-Vedele F., et al.. (1999) Molecular and functional regulation of two NO3– uptake systems by N- and C-status of Arabidopsis plants. Plant J. 18: 509–519.
- Leran S., Varala K., Boyer J.C., Chiurazzi M., Crawford N., Daniel-Vedele F., et al.. (2014) A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 19: 5–9.
- Lezhneva L., Kiba T., Feria-Bourrellier A.B., Lafouge F., Boutet-Mercey S., Zoufan P., et al.. (2014) The Arabidopsis nitrate transporter NRT2.5 plays a role in nitrate acquisition and remobilization in nitrogen-starved plants. Plant J. 80: 230–241.
- Liang G., Ai Q., Yu D.Q. (2015) Uncovering miRNAs involved in crosstalk between nutrient deficiencies in Arabidopsis. Sci. Rep. 5: 11813.
- Lima J.E., Kojima S., Takahashi H., von Wiren N. (2010) Ammonium triggers lateral root branching in Arabidopsis in an AMMONIUM TRANSPORTER1;3-dependent manner. Plant Cell 22: 3621–3633.
- Lin S.H., Kuo H.F., Canivenc G., Lin C.S., Lepetit M., Hsu P.K., et al.. (2008) Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 20: 2514–2528.
- Liu K.H., Tsay Y.F. (2003) Switching between the two action modes of the dual-affinity nitrate transporter CHL1 by phosphorylation. EMBO J. 22: 1005–1013.
- Liu X.Q., Huang D.M., Tao J.Y., Miller A.J., Fan X.R., Xu G.H. (2014) Identification and functional assay of the interaction motifs in the partner protein OsNAR2.1 of the two-component system for high-affinity nitrate transport. New Phytol. 204: 74–80.
- Lopez-Bucio J., Cruz-Ramirez A., Herrera-Estrella L. (2003) The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 6: 280–287.
- Loque D., Yuan L., Kojima S., Gojon A., Wirth J., Gazzarrini S., et al.. (2006) Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 48: 522–534.
- Ludewig U., Neuhduser B., Dynowski M. (2007) Molecular mechanisms of ammonium transport and accumulation in plants. FEBS Lett. 581: 2301–2308.
- Ma J.F., Yamaji N. (2015) A cooperative system of silicon transport in plants. Trends Plant Sci. 20: 435–442.
- Ma W.Y., Li J.J., Qu B.Y., He X., Zhao X.Q., Li B., et al.. (2014) Auxin biosynthetic gene TAR2 is involved in low nitrogen-mediated reprogramming of root architecture in Arabidopsis. Plant J. 78: 70–79.
- Ma Q., Tang R.J., Zheng X.J., Wang S.M., Luan S. (2015) The calcium sensor CBL7 modulates plant responses to low nitrate in Arabidopsis. Biochem. Biophys. Res. Commun. 468: 59–65.
- Marchive C., Roudier F., Castaings L., Brehaut V., Blondet E., Colot V., et al.. (2013) Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 4: 1713.
- Miller A.J., Cramer M.D. (2004) Root nitrogen acquisition and assimilation. Plant Soil 27: 1–36.
- Miwa K., Fujiwara T. (2010) Boron transport in plants: co-ordinated regulation of transporters. Ann. Bot. 105: 1103–1108.
- Mueller N.D., Gerber J.S., Johnston M., Ray D.K., Ramankutty N., Foley J.A. (2012) Closing yield gaps through nutrient and water management. Nature 490: 254–257.
- Munos S., Cazettes C., Fizames C., Gaymard F., Tillard P., Lepetit M., et al.. (2004) Transcript profiling in the chl1‐5 mutant of Arabidopsis reveals a role of the nitrate transporter NRT1.1 in the regulation of another nitrate transporter, NRT2.1. Plant Cell 16: 2433–2447.
- Nacry P., Bouguyon E., Gojon A. (2013) Nitrogen acquisition by roots: physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 370: 1–29.
- Nguyen G.N., Rothstein S.J., Spangenberg G., Kant S. (2015) Role of microRNAs involved in plant response to nitrogen and phosphorous limiting conditions. Front. Plant Sci. 6: 629.
- Pant B.D., Musialak-Lange M., Nuc P., May P., Buhtz A., Kehr J., et al.. (2009) Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 150: 1541–1555.
- Parker J.L., Newstead S. (2014) Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1. Nature 507: 68–72.
- Pellizzaro A., Clochard T., Planchet E., Limami A.M., Morere-Le Paven M.C. (2015) Identification and molecular characterization of Medicago truncatula NRT2 and NAR2 families. Physiol. Plant. 154: 256–269.
- Postma J.A., Dathe A., Lynch J.P. (2014) The optimal lateral root branching density for maize depends on nitrogen and phosphorus availability. Plant Physiol. 166: 590–602.
- Rawat S.R., Silim S.N., Kronzucker H.J., Siddiqi M.Y., Glass A.D. (1999) AtAMT1 gene expression and NH4+ uptake in roots of Arabidopsis thaliana: evidence for regulation by root glutamine levels. Plant J. 19: 143–152.
- Riveras E., Alvarez J.M., Vidal E.A., Oses C., Vega A., Gutierrez R.A. (2015) The calcium ion is a second messenger in the nitrate signaling pathway of Arabidopsis. Plant Physiol. 169: 1397–1404.
- Rubin G., Tohge T., Matsuda F., Saito K., Scheible W.R. (2009) Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 21: 3567–3584.
- Ruffel S., Krouk G., Ristova D., Shasha D., Birnbaum K.D., Coruzzi G.M. (2011) Nitrogen economics of root foraging: transitive closure of the nitrate–cytokinin relay and distinct systemic signaling for N supply vs. demand. Proc. Natl. Acad. Sci. USA 108: 18524–18529.
- Sonoda Y., Ikeda A., Yamaya T., Yamaguchi J. (2004) Feedback regulation of the ammonium transporter gene family AMT1 by glutamine in rice. Plant Cell Physiol. 45: S98.
- Sun J., Bankston J.R., Payandeh J., Hinds T.R., Zagotta W.N., Zheng N. (2014) Crystal structure of the plant dual-affinity nitrate transporter NRT1.1. Nature 507: 73–77.
- Tabata R., Sumida K., Yoshii T., Ohyama K., Shinohara H., Matsubayashi Y. (2014) Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 346: 343–346.
- Takano J., Tanaka M., Toyoda A., Miwa K., Kasai K., Fuji K., et al.. (2010) Polar localization and degradation of Arabidopsis boron transporters through distinct trafficking pathways. Proc. Natl. Acad. Sci. USA 107: 5220–5225.
- Terasaka K., Blakeslee J.J., Titapiwatanakun B., Peer W.A., Bandyopadhyay A., Makam S.N., et al.. (2005) PGP4, an ATP binding cassette P-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots. Plant Cell 17: 2922–2939.
- Walch-Liu P., Forde B.G. (2008) Nitrate signalling mediated by the NRT1.1 nitrate transporter antagonises L-glutamate-induced changes in root architecture. Plant J. 54: 820–828.
- Wang Q.L., Zhao Y.Y., Luo W.X., Li R.L., He Q.H., Fang X.H., et al.. (2013) Single-particle analysis reveals shutoff control of the Arabidopsis ammonium transporter AMT1;3 by clustering and internalization. Proc. Natl. Acad. Sci. USA 110: 13204–13209.
- Werdin-Pfisterer N.R., Kielland K., Boone R.D. (2012) Buried organic horizons represent amino acid reservoirs in boreal forest soils. Soil Biol. Biochem. 55: 122–131.
- Wirth J., Chopin F., Santoni V., Viennois G., Tillard P., Krapp A., et al.. (2007) Regulation of root nitrate uptake at the NRT2.1 protein level in Arabidopsis thaliana. J. Biol. Chem. 282: 23541–23552.
- Xu G.H., Fan X.R., Miller A.J. (2012) Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 63: 153–182.
- Yan M., Fan X.R., Feng H.M., Miller A.J., Shen Q.R., Xu G.H. (2011) Rice OsNAR2.1 interacts with OsNRT2.1, OsNRT2.2 and OsNRT2.3a nitrate transporters to provide uptake over high and low concentration ranges. Plant Cell Environ. 34: 1360–1372.
- Yang X.Q., Lee S., So J.H., Dharmasiri S., Dharmasiri N., Ge L., et al.. (2004) The IAA1 protein is encoded by AXR5 and is a substrate of SCFTIR1. Plant J. 40: 772–782.
- Yong Z.H., Kotur Z., Glass A.D.M. (2010) Characterization of an intact two-component high-affinity nitrate transporter from Arabidopsis roots. Plant J. 63: 739–748.
- Yuan L., Loque D., Kojima S., Rauch S., Ishiyama K., Inoue E., et al.. (2007a) The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 19: 2636–2652.
- Yuan L.X., Gu R.L., Xuan Y.H., Smith-Valle E., Loque D., Frommer W.B., et al.. (2013) Allosteric regulation of transport activity by heterotrimerization of Arabidopsis ammonium transporter complexes in vivo. Plant Cell 25: 974–984.
- Yuan L.X., Loque D., Ye F.H., Frommer W.B., von Wiren N. (2007b) Nitrogen-dependent posttranscriptional regulation of the ammonium transporter AtAMT1;1. Plant Physiol. 143: 732–744.
- Zhao M., Ding H., Zhu J.K., Zhang F., Li W.X. (2011) Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 190: 906–915.