Combination antiretroviral therapy (ART) suppresses viral replication to undetectable levels, reduces morbidity and mortality, and improves the quality of life for individuals living with human immunodeficiency virus (HIV) []. However, ART cannot cure HIV infection because it is unable to eliminate latently HIV-infected cells. Hence, new antiviral drugs are needed to target latent HIV proviruses in the absence of active replication.
The antiviral drug ABX464 is an oral, first-in-class compound that inhibits HIV replication in vitro through a new mechanism of action by binding the cap binding complex, inhibiting the viral protein Rev, and altering HIV mRNA biogenesis []. ABX464 has demonstrated safety and profound antiinflammatory activity in preclinical trials and in a completed phase 2a proof-of-concept study to treat lesions in ulcerative colitis (ABIVAX, data in file). Additionally, ABX464 is the first HIV antiviral that in transgenic humanized mice induced a long-term control of the viral load after treatment arrest []. In the phase 2a clinical trial ABX464-004 (NTC02735863), ABX464 demonstrated a decrease in total HIV-DNA in peripheral CD4 + T cells in 2 of 2 participants treated with 50 mg of ABX464 daily for 28 days and in 6 of 14 treated with 150 mg of ABX464 daily for 28 days, while no changes were observed in the placebo group (n = 4) []. Furthermore, in the recent phase 2a clinical trial ABX464-005 (NCT02990325), ABX464 showed a reduction of total HIV-DNA in peripheral CD4 + T cells after 28 days of combination treatment with ART []. These results suggest that ABX464 is a promising new antiviral drug that may affect HIV RNA biogenesis and eliminate HIV-infected cells in vivo. However, none of these clinical trials evaluated the specific effect of this antiviral drug on intact HIV DNA or HIV transcription in vivo. In this study, we quantified total and intact HIV DNA and characterized the HIV transcription profile before and after ABX464 therapy in available samples from the ABX464-005 (NCT02990325) study.
METHODS
Study Design
The study is a follow-up to the ABIVAX-005 clinical trial (NCT02990325). In this trial, 24 ART-suppressed individuals living with HIV and 12 individuals not living with HIV received daily oral doses of the antiviral drug ABX464 (Abivax, Paris, France). Eleven individuals living with HIV were treated with 150 mg for 4 weeks, and 13 individuals living with HIV and 12 individuals not individuals living with HIV were treated with 50 mg for 12 weeks.
Ethics Statement
The Spanish Agency of Medicines and Medical Products approved the trial. Each participant provided written informed consent.
Samples
Cryopreserved peripheral blood mononuclear cells (PBMCs) from weeks 0, 4, and 8 were available from 9 ART-suppressed cisgender men living with HIV treated with 150 mg daily orally for 4 weeks. CD4+ T cells were purified by negative selection (Miltenyi) and immediately frozen as dry pellets. Nucleic acids were extracted using TRI Reagent [] and used to quantify HIV DNA and HIV transcripts.
Total and Intact HIV DNA
Total HIV DNA (long terminal repeat [LTR] or Gag regions to avoid undetectable values due to sequence mismatch) was quantified in duplicate using droplet digital polymerase chain reaction (ddPCR) []. Intact, hypermutated/3’defective and 5’defective HIV DNA (Packaging signal [Ψ] and Env [Rev Response Element) were quantified in 8 replicates using the intact proviral DNA assay by ddPCR [] (Figure 1A).
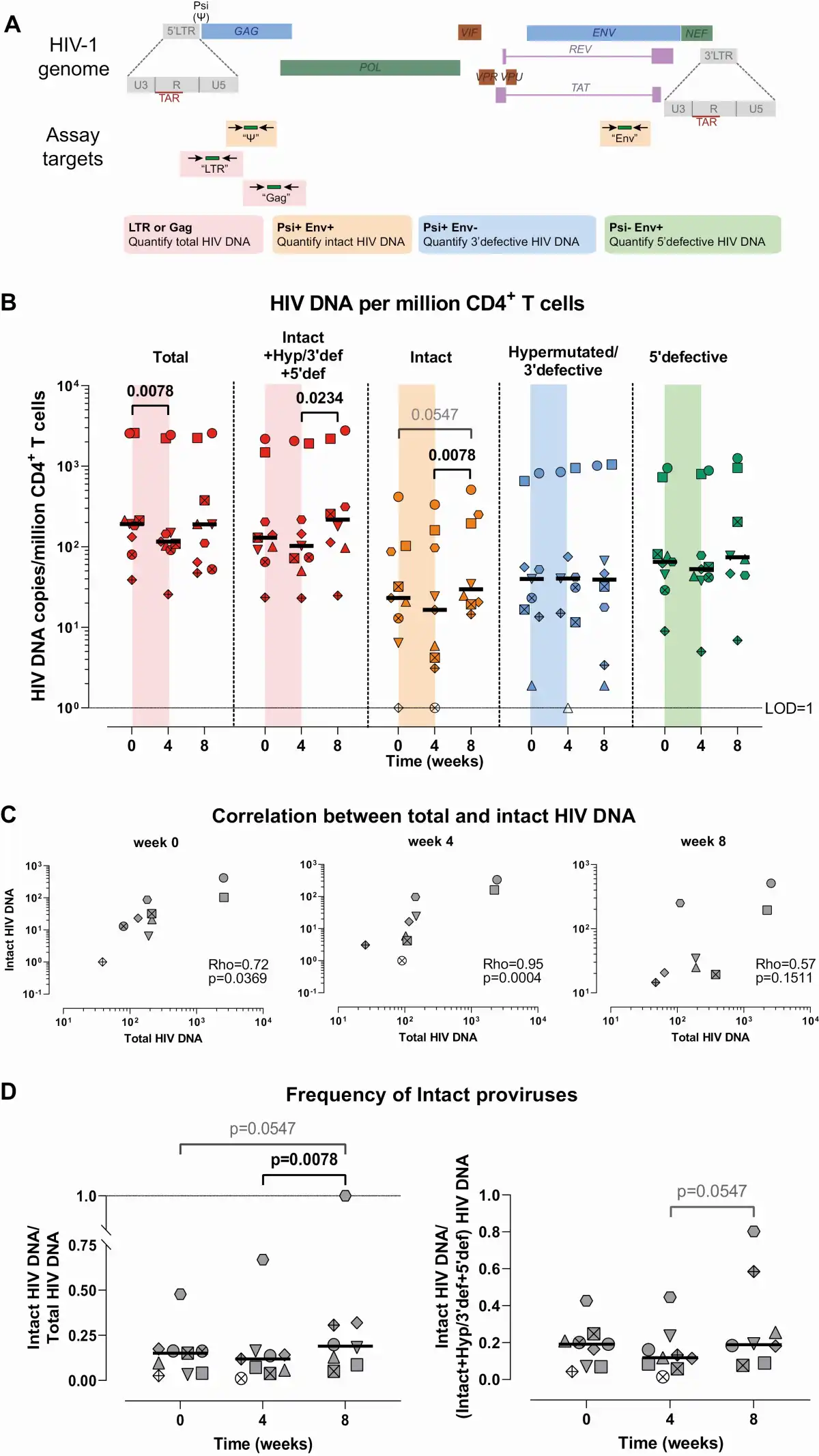
Figure 1
Effect of ABX464 on the HIV reservoir. A, Diagram of the assays used to quantify total, intact, 3’defective, and 5’defective HIV DNA. B, Dynamics of total, intact, 3’defective, and 5’defective HIV DNA. C, Spearman correlations between total (long terminal repeat [LTR] or Gag) and intact proviral DNA assay (IPDA) HIV DNA. D, Relative frequency of IPDA over total (LTR or Gag) HIV DNA and of IPDA over the sum of hypermutated/3’defective, 5’defective, and intact HIV DNA (values ≤1 were censored to 1). Each color represents a different assay; each symbol/shape represents a different individual from the ABX464-005 study; bars represent medians. Determinations below the limit of detection are represented as open dots. Abbreviation: HIV, human immunodeficiency virus.
HIV Transcript Levels
Cell-associated HIV transcripts (initiated [TAR], 5’elongated [R-U5-preGag], unspliced [Pol], polyadenylated [PolyA], and multiply-spliced [Tat-Rev]) were quantified in duplicate using reverse-transcription-ddPCR [] (Figure 2A).
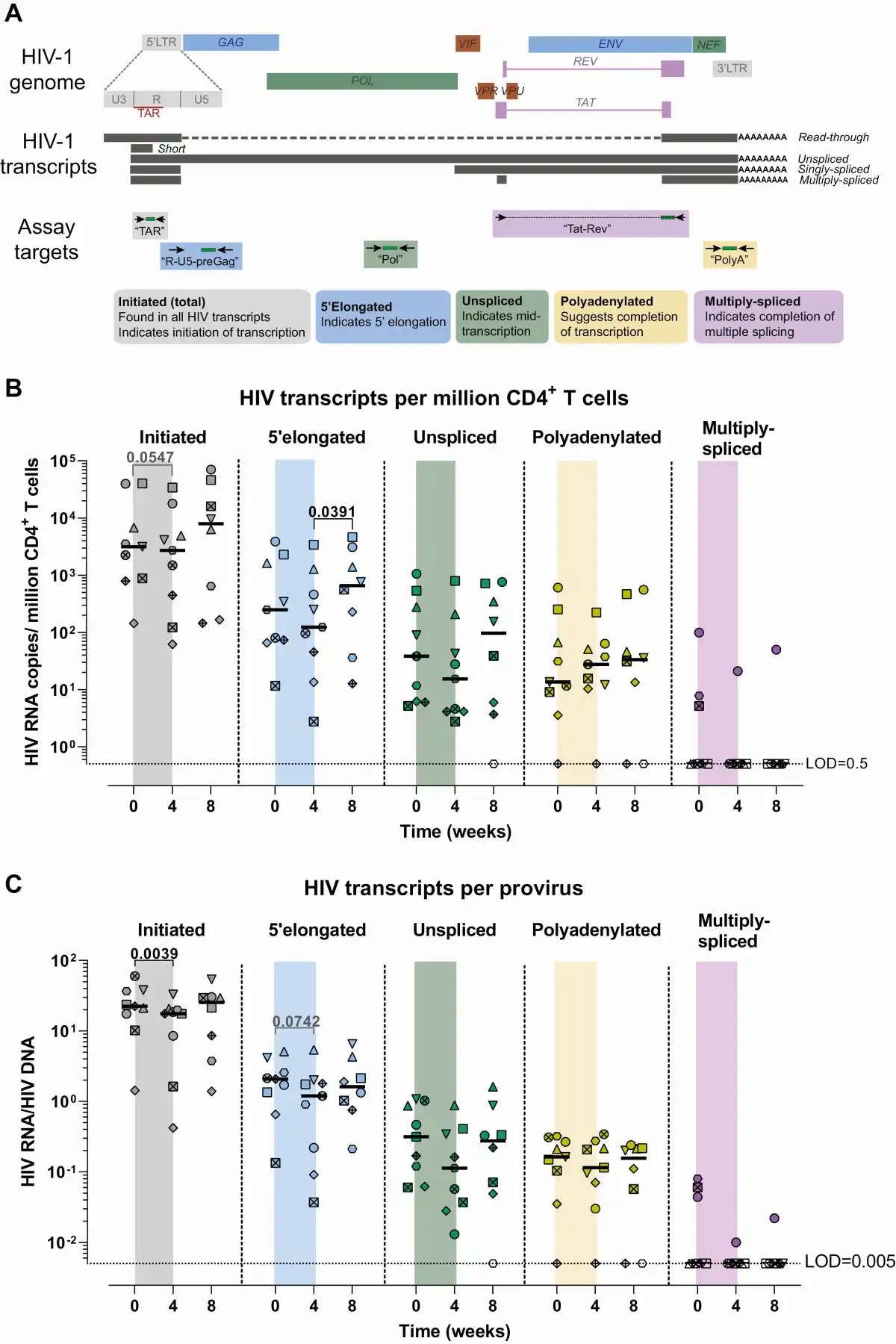
Figure 2
Effect of ABX464 on HIV transcription. A, Diagram of the assays used to characterize the HIV transcription profile. B, Dynamics of each HIV transcript per million CD4+ T cells. C, Dynamics of each HIV transcript per provirus (HIV RNA-to-HIV DNA). Each color represents a different assay; each symbol/shape represents a different individual; bars represent medians. Determinations below the limit of detection are represented as open dots. Abbreviation: HIV, human immunodeficiency virus.
Statistics
Wells without positive droplets were assigned the limit of detection of 1 copy/replicate. Longitudinal changes were evaluated using the Wilcoxon signed-rank test, and correlations were assessed using the Spearman test. GraphPad Prism version 7.0 was used for all statistical analyses.
RESULTS
Characteristics of the Study Participants
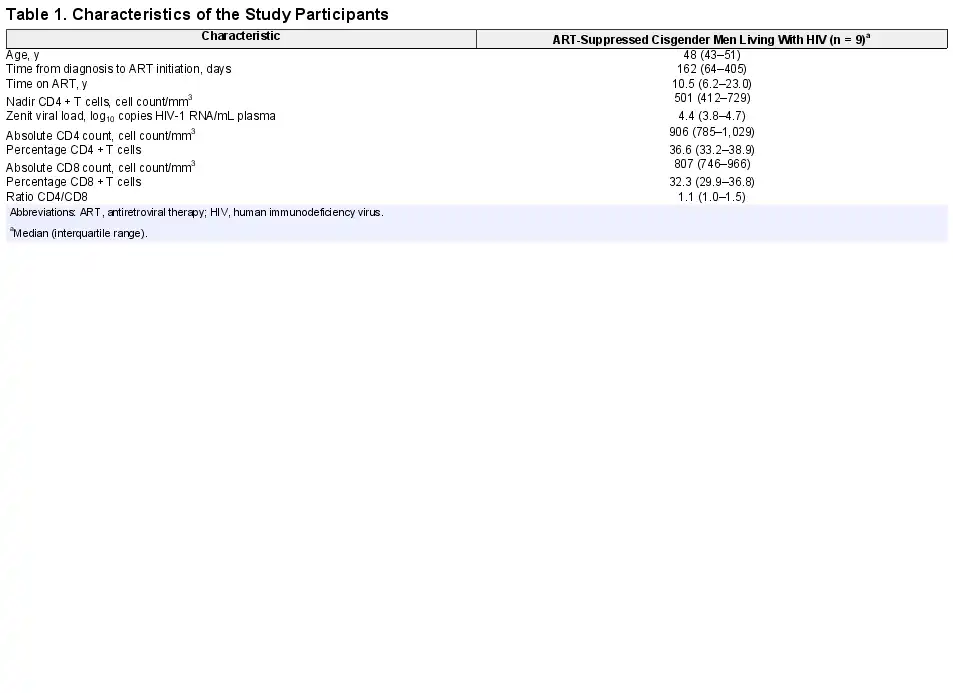
Cryopreserved PBMCs from 9 ART-suppressed cisgender men living with HIV treated with 150 mg of ABX464 were available from weeks 0, 4, and 8 to measure HIV DNA (total and intact) and HIV transcript levels. Baseline characteristics of the study participants are summarized in Table 1. Briefly, the median (interquartile range) time from diagnosis to ART initiation was 162 days (64–405), and the median time on ART was 10.5 years (6.2–23.0).
ABX464 Decreases Total HIV DNA and May Decrease Intact HIV DNA
We quantified total, intact, 3’ defective, and 5’ defective HIV DNA (Figure 1A) at weeks 0 (before ABX464 treatment initiation), 4 (at ABX464 treatment discontinuation), and 8 (4 weeks after ABX464 discontinuation). Total HIV DNA decreased significantly on ABX464 (P = .0078, median [week 4/week 0] = 0.8). Intact and 5’ defective HIV DNA showed a similar trend (P = not significant [n.s.], median [week 4/week 0] = 0.8 for both), while no such trend was observed for hypermutated/3’defective HIV DNA. After discontinuation of ABX464, however, the intact HIV DNA increased significantly from week 4 to week 8 (P = .0078, median [week 8/week 4] = 1.4), and a trend was observed toward smaller increases in total (median [week 8/week 4] = 1.2) and 5’defective (median [week 8/week 4] = 1.4) HIV DNA (Figure 1B). These data suggest that ABX464 treatment decreases the total HIV reservoir and may also decrease the intact HIV reservoir when it is administered in combination with ART. Hence, ABX464 may act as an ART intensifier or improve ART effect.
Since we observed differences in the increase of total and intact HIV DNA after ABX464 discontinuation, we sought to determine whether the correlation between total and intact was affected by ABX464. Total and intact HIV DNA correlated strongly at weeks 0 (Rho = 0.72, P = .037) and 4 (Rho = 0.95, P = .0004), while at week 8, the Rho value was less and the P value was no longer statistically significant (Rho = 0.57, P = .15; Figure 1C).
Furthermore, the relative ratio of intact to total (LTR or Gag) proviruses increased after treatment discontinuation (P = .0078 for week 4 vs week 8; P = .055 for week 0 vs week 8), and a similar trend was observed for the ratio of intact proviruses to the sum of hypermutated/3’defective, 5’defective, and intact proviruses (P = .055 for week 4 vs week 8; Figure 1D). These results suggest that while ABX464 tended to decrease total and intact HIV DNA, the intact HIV DNA tended to increase more after treatment discontinuation.
ABX464 Decreases HIV Transcriptional Initiation and May Decrease HIV Elongation
To determine the effect of ABX464 treatment on HIV transcription during ART, we quantified HIV initiated (total), 5’elongated, unspliced, polyadenylated, and multiply-spliced HIV transcripts (Figure 2A) at weeks 0, 4, and 8. Each HIV transcript was expressed as copies per million peripheral CD4+ T cells (Figure 2B) and also (given the changes in HIV DNA) normalized to copies per provirus (HIV RNA-to-HIV DNA ratio) (Figure 2C). Administration of ABX464 was associated with a significant decrease in initiated HIV transcripts per provirus (P = .0039, median [week 4/week 0] = 0.5), a trend toward a decrease in 5’elongated HIV transcripts per provirus (P = .074, median fold change = 0.5), and a lower median level of unspliced HIV RNA per provirus (P = n.s., median fold change = 0.6), while no decrease was observed in polyadenylated or multiply-spliced HIV transcripts. However, 5’elongated HIV RNA per million CD4+ T cells increased significantly (P = .04, fold change = 1.4) after ABX464 discontinuation. These data suggest that ABX464 treatment decreases HIV transcription when it is administered in combination with ART. Hence, ABX464 may act directly or indirectly as a latency silencing agent.
DISCUSSION
We characterized the levels of total and intact HIV DNA and the levels of various HIV transcripts before and after administration of the antiviral drug ABX464 in combination with ART. We are the first to evaluate in vivo the specific effect of ABX464 on the intact HIV reservoir and the HIV transcription profile. We detected a significant decrease in total HIV DNA and a similar trend in intact HIV DNA after 4 weeks of ABX464 treatment. The changes in total HIV DNA agreement with those observed in ABX464-004 [] and the parental study ABX464-005 []. It is unclear if the decrease in HIV DNA represents selective cell migration/homing, decreased proliferation, or death of infected cells vs another process. Regarding the mechanisms of action of ABX464 reported by Vautrin et al [], we hypothesize that by altering HIV splicing, this drug may induce the production of aberrant HIV transcripts that might be recognized as viral antigens, facilitating the elimination of infected cells. However, the HIV DNA increased again after ABX464 discontinuation, further suggesting that the changes were due to ABX464 and are reversible (as for the effects of ART). Furthermore, we observed an increase in the relative frequency of intact proviruses after ABX464 discontinuation, which may reflect a specific proliferation of cells harboring intact proviruses.
We also observed a significant decrease in initiated (total) HIV transcripts per provirus and a trend toward a decrease in 5’elongated HIV transcripts per provirus. As for HIV DNA, these decreases reversed after ABX464 discontinuation. However, we observed no statistically significant change in multiply-spliced HIV transcripts, in contrast to a previous study in which in vitro treatment with ABX464 enhanced alternative splicing of HIV RNA in infected PBMCs from 6 healthy individuals []. This discrepancy may reflect specific differences in the effect of ABX464 in vivo vs in vitro or the fact that we only measured multiply-spliced HIV Tat-Rev transcripts and so may have missed changes in multiply-spliced Nef or singly or alternatively-spliced HIV transcripts. Moreover, the very low baseline levels of multiply-spliced Tat-Rev and the high proportion of samples with no detectable Tat-Rev might have interfered with the ability to detect changes in these transcripts.
ABX464 treatment in vitro increased the splicing of the long noncoding RNA lncRNA0599-205, which encodes for the microRNA miR-124, increasing its expression []. Furthermore, the parental study ABX464-005 showed an increase in miR-124 during ABX464 administration []. miR-124 is a critical modulator of immunity and inflammation and acts as an antiinflammatory microRNA by inhibiting the production of proinflammatory cytokines []. In this regard, the parental study ABX464-005 showed a decrease in inflammation markers in plasma and a decrease in the percent of activated CD4+ T cells in blood (unpublished data). Since T-cell activation stimulates HIV transcription, it is possible that an ABX464-mediated decrease in T-cell activation contributes to the decrease in HIV transcriptional initiation observed in the present substudy [, ]. Alternatively, an ABX464-mediated decrease in HIV transcription could also contribute to a decrease in inflammation, or there could be bidirectional effects.
Limitations of the study should be acknowledged. First, samples were only available from 9 of 11 trial participants treated with 150 mg of ABX464. Second, most of the participants presented with lower HIV levels, which may have affected the detection of changes in total and intact HIV DNA and HIV transcripts. While we detected a statistically significant decrease in the expression of initiated (total) HIV transcripts and a trend toward a decrease in 5’elongated and unspliced HIV transcripts after ABX464 administration, it is difficult to exclude a small effect on polyadenylated and multiply-spliced transcripts due to the number of undetectable determinations.
In conclusion, our findings have important implications for strategies aimed at HIV cure. ABX464 administration in vivo had a dual effect of decreasing HIV DNA (total and possibly intact) as well as HIV transcription. Therefore, this antiviral drug and similar agents may play a role in strategies aimed at reducing the HIV reservoir and/or preventing reactivation from latently infected cells. Future studies should investigate the specific mechanism(s) that reduce HIV DNA and RNA in vivo after ABX464 treatment in combination with ART.
Notes
Author Contributions. S. M.-L., J. M.-P., and S. A. Y. designed the substudy. S. M. L. performed experiments, analyzed data, and (with S. A. Y.) wrote the manuscript. S. B. S. performed experiments and analyzed data. J. M.-P. designed the clinical trial, provided samples, and assisted with manuscript preparation. S. A. Y. and J. K. W. supervised the work. All authors edited the manuscript. All authors read and accepted the final manuscript.
Acknowledgments. The authors thank the study participants who made this study possible.
Disclaimer. Abivax sponsored the clinical study from which these samples were derived and contributed to the design of the clinical trial. The funders had no role in data collection and analysis, decision to publish, or preparation of the manuscript.
Financial support. This work was supported by the University of California–San Francisco Gladstone Center for AIDS Research (P30A1027763 to S. M.-L.]); the National Institute of Allergy and Infectious Diseases at the National Institutes of Health (NIH; 1R01AI132128 to S. A. Y. and J. K. W.; R56AI116342, R33AI116218, R56AI091573, and U19AI096109 to J. K. W.); the National Institute of Diabetes and Digestive and Kidney Diseases at the NIH (1R01DK108349 and 1R01DK120387 to S. A. Y.); the US Department of Veterans Affairs (IK2 CX000520 to S. A. Y.; I01 BX000192 to J. K. W.); and the American Foundation for AIDS Research Institute for HIV Cure Research (109301 to S. A. Y. and J. K. W.]). J.M.-P. is supported by grant PID2019-109870RB-I00 from the Spanish Ministry of Science and Innovation and by RETIC RD16/0025/0041 (cofunded by the European Regional Development Fund/European Social Fund).
Potential conflicts of interest. J. M.-P. received grant support and consultancy fees from Abivax. J. K. W. reports grants/support from the US Department of Veteran Affairs and the NIH outside the submitted work. J. M.-P reports research grants paid to their institution from Grifols during the conduct of the study; unrestricted research grants to their institution from MSD, ViiV Healthcare, Gilead Sciences, and AstraZeneca; consulting fees from MSD; and payment for educational activities from Janssen and Gilead, all outside the submitted work. S. A. Y. reports salary support from US Department of Veterans Affairs and grants from the NIH and the American Foundation for AIDS Research during the conduct of the study, all outside the submitted work. All other authors report no potential conflicts.
All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References
- 1. Chun TW, Stuyver L, Mizell SB, et al Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc Natl Acad Sci U S A1997; 94:13193–7.
- 2. Wong JK, Hezareh M, Günthard HF, et al Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science1997; 278:1291–5.
- 3. Finzi D, Hermankova M, Pierson T, et al Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science1997; 278:1295–300.
- 4. Vautrin A, Manchon L, Garcel A, et al Both anti-inflammatory and antiviral properties of novel drug candidate ABX464 are mediated by modulation of RNA splicing. Sci Rep2019; 9:792.
- 5. Tazi PJ. Durable control of viral rebound in humanized mice by ABX464 targeting rev functions. In: Conference on Retroviruses and Opportunistic Infections, CROI 2015, Seattle. 2015: Oral presentation.
- 6. Rutsaert S, Steens JM, Gineste P, et al Safety, tolerability and impact on viral reservoirs of the addition to antiretroviral therapy of ABX464, an investigational antiviral drug, in individuals living with HIV-1: a phase IIa randomised controlled study. J Virus Erad2019; 5:10–22.
- 7. Steens J-M, Cranston R, Martinez-Picado J, et al Oral ABX464 reduces the HIV DNA reservoir in CD4+ peripheral blood T cells. J Virus Eradicat2017; 3:1–59.
- 8. Yukl SA, Kaiser P, Kim P, et al HIV latency in isolated patient CD4+T cells may be due to blocks in HIV transcriptional elongation, completion, and splicing. Sci Transl Med2018; 10:1–16.
- 9. Morón-López S, Puertas MC, Gálvez C, et al Sensitive quantification of the HIV-1 reservoir in gut-associated lymphoid tissue. PLoS One2017; 12:e0175899.
- 10. Bruner KM, Wang Z, Simonetti FR, et al A quantitative approach for measuring the reservoir of latent HIV-1 proviruses. Nature2019; 566:120–5.
- 11. Sun Y, Li Q, Gui H, et al MicroRNA-124 mediates the cholinergic anti-inflammatory action through inhibiting the production of pro-inflammatory cytokines. Cell Res2013; 23:1270–83.
- 12. Cheadle C, Fan J, Cho-Chung YS, et al Control of gene expression during T cell activation: alternate regulation of mRNA transcription and mRNA stability. BMC Genomics2005; 6:75.
- 13. Morón-López S, Gómez-Mora E, Salgado M, et al Short-term treatment with interferon alfa diminishes expression of HIV-1 and reduces CD4 + T-cell activation in patients coinfected with HIV and hepatitis C virus and receiving antiretroviral therapy. J Infect Dis2015; 213:1008–12.