Dicarboxylic acids as the missing fourth nutrient
Medium- and long-chain dicarboxylic acids (DAs) are present in higher plants and animals, deriving from the ω-oxidation of fatty acids (). In higher plants, DAs are components of natural protective polymers, such as cutin and suberin, a support biopolyester that waterproofs the leaves and fruits, regulating the flow of nutrients and minimizing the harmful impact of pathogens (). They are β-oxidized in specialized plant peroxisomes, called glyoxysomes, where the glyoxylate cycle, whose intermediates derive from the degradation of reserve or structural lipids, takes place ().
The paradigm of nutrient composition acknowledges only three energy substrates: glucose, amino acids, and fatty acids. In this issue of the JCI, Goetzman et al., () confirmed and magnified the evidence of previous investigations establishing the presence and metabolic importance of a fourth nutrient, DAs. While metabolizable DAs are classified as medium-chain fatty acids, they possess two terminal carboxylic groups that permit the formation of dicarboxylic salts, making them easily soluble in water and promptly available as energy. Contrary to typical dietary fatty acids, DAs do not require transport within the circulatory stream, similar to triglycerides, which, as they are insoluble in water, must be incorporated into chylomicrons or lipoproteins such as very-low-density lipoproteins (VLDL) or low-density lipoproteins (LDL). DAs can be immediately β-oxidized, giving rise to acetyl-CoA and succinyl-CoA. Some diseases, such as type 2 diabetes (T2D), result in an inability to rapidly adjust energy substrate utilization and show shortage of glycogen depots in both the liver and skeletal muscle due to impaired glycogen synthesis. Instead, DAs are completely oxidized in T2D in the liver as well as in the muscle and they restore glycogen depots and improve glycemic control (). Another important characteristic of DAs is their ability to access the mitochondria without the carnitine shuttle, a feature that facilitates complete oxidation ().
DAs spare glucose utilization and increase glycogen stores in T2D in animal models and in humans (–). In fact, 10 g or 23 g of a DA with 10 carbon atoms, such as sebacic acid, added to a 450 kcal meal containing 75 g of glucose but without lipids drastically reduced the plasma glucose peak in people with T2D (). The circulating levels of insulin in response to the meal were reduced with both amounts of sebacic acid and the rate at which glucose appeared in the circulation after eating was decreased by about 18% (P < 0.05) ().
DC12 improves insulin resistance and increases energy expenditure
DAs represent a highly performing energy substrate during physical exercise. While metabolically healthy skeletal muscle can switch easily between glucose and fat oxidation in response to homeostatic signals, the skeletal muscle from individuals with T2D or obesity shows a great reduction in this metabolic flexibility (). Dodecanedioic acid (DC12), a 12-carbon atom straight-chain DA, increases skeletal muscle glycogen stores in people with T2D and might postpone the development of fatigue and increase exercise time ().
Goetzman et al., () fed mice with diets containing various fat combinations during a nine-week course. Unlike most dietary fats, DAs could not be stored, but, rather, were completely oxidized. Consequently, DA intake did not increase fat mass, despite consumption of 33% of calories as fat (Figure 1). In an era where obesity has become a pandemic (), DAs might represent a way to reduce the obesity burden in association with antiobesity medications and minimally invasive surgery. Moreover, Goetzman and colleagues () demonstrated that DAs, in particular DC12, increased energy expenditure.
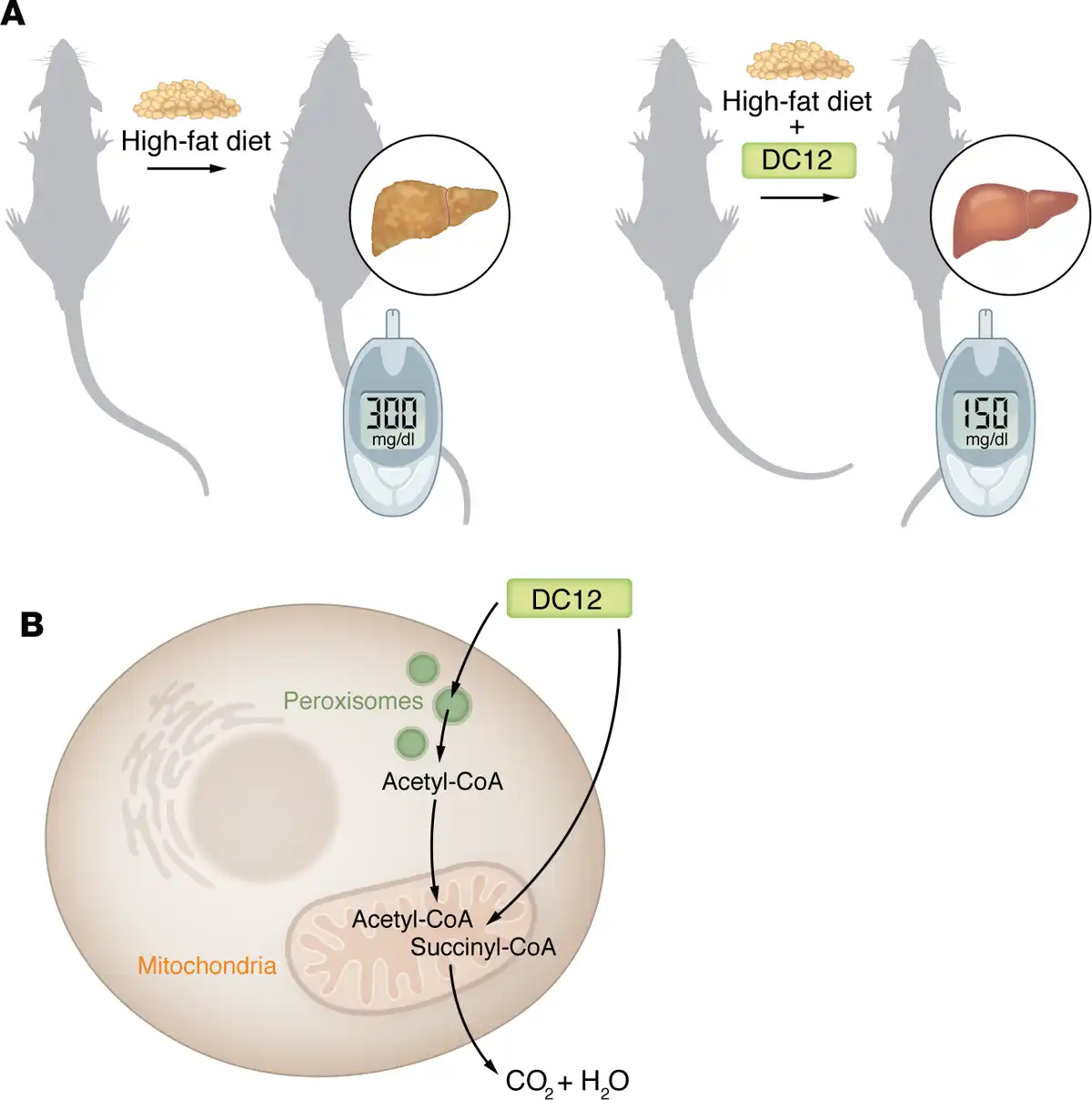
Figure 1
DC12 serves as an alternative source of dietary fat.
(A) Mice fed a high-fat diet very rich in saturated fatty acids develop obesity and hepatic steatosis and show enhanced circulated levels of glucose and insulin after intraperitoneal glucose administration. In contrast, the addition of 10% DC12 prevents such metabolic disorders. (B) DC12 undergoes β-oxidation in the mitochondria and peroxisomes to form the end products acetyl-CoA and succinyl-CoA. While DC12 is completely oxidized to CO2 and H2O in the mitochondria, acetyl-CoA produced in peroxisomes is transported into the mitochondria where it is completely oxidized.
Interestingly, Goetzman et al. () proved that excess of succinate formation from DA oxidation increased protein succinylation also in white adipose tissue, promoting a process called “beiging” by binding the adipocyte surface through the succinate receptor GPR91. This is a very interesting observation. In fact, it is known that white adipose tissue can undergo the process of browning in response to a variety of stimuli inducing or enhancing the expression of thermogenes that are typically associated with brown fat. White adipose cells become beige or brite cells and are characterized by higher energy expenditure and reduced fat storage ().
Therefore, a high-fat diet substituted with DC12 increased metabolic rate and prevented obesity in mice, a finding that represents a strategic nutrient approach to slow the progression of the global obesity epidemic. Importantly, mice administered DC12 maintained a normal insulin sensitivity without an increase in blood glucose.
T2D is preceded by a latent period, known as prediabetes, which is caused by peripheral insulin resistance and a compensatory increase in circulating levels of insulin; almost one in three individuals in the United States in 2020 had insulin resistance (). When pancreatic β cells become unable to compensate the insulin resistance state and insulin secretion fails, overt T2D manifests. A high-fat diet is associated with obesity, whole-body insulin resistance, hyperlipidemia, and glucose intolerance (). DC12 can overcome such metabolic derangement, preserving the pancreatic insulin secretion machinery and avoiding insulin resistance associated glucose intolerance.
Further, dodecanedioic acid and the shorter DA with 10 carbon atoms, sebacic acid, were proven to improve glucose disposal in humans (–).
Another important advantage of dietary DC12 over fatty acids that was highlighted in Goetzman et al. () is its storage; DC12 was not stored intracellularly and thus it did not promote fat deposition. In other words, DAs are completely oxidized either in mitochondria or in peroxisomes (Figure 1).
Dodecanedioic acids counteract the effects of high-fat diet
Supplementation of a 60% high-fat diet with DC12 prevented liver triglyceride accumulation, a feature present in the metabolic-associated fatty liver disease (MAFLD). MAFLD () is a pathologic condition, associating hepatic steatosis with T2D and/or obesity and/or metabolic dysregulation (), with a high prevalence in adults ranging worldwide between 17% and 51% and with the potential to progress to liver cirrhosis and/or hepatocellular carcinoma ().
Conclusions and implications
An important question remains open, whether DC12 prevents or reverses liver steatosis alone or if it is also effective in metabolic dysfunction-associated steatohepatitis (MASH). Although the global prevalence of MASH is 5.27% (), far lower than the prevalence of MAFLD, its clinical picture and evolution are much more severe. Therefore, this topic should be specifically addressed in the future given the scarcity of available therapies (, ). It will be critical to perform clinical trials to prove the translational safety and efficacy of DC12.
DAs in general, and DC12 in particular, are safe nutrients that are completely oxidized. They increase energy expenditure without accumulation in adipose tissue, thereby preventing weight gain. Their intake reduces the effects of dietary triglycerides that cause insulin resistance, hyperlipidemia, and liver steatosis. DAs represent the fourth known nutrient — in addition to glucose, amino acids, and fatty acids — known to improve metabolic derangement associated with a high-fat diet, typical of Westernized societies.
- 1. Mingrone G, Castagneto M. Medium-chain, even-numbered dicarboxylic acids as novel energy substrates: an update Nutr Rev. 2006;64(10 pt 1):449–456
- 2. Goetzman E, et al Dietary dicarboxylic acids provide a non-storable alternative fat source that protects mice against obesity J Clin Invest. 2024;134(12)e174186
- 3. Iaconelli A, et al Effect of oral sebacic acid on postprandial glycemia, insulinemia, and glucose rate of appearance in type 2 diabetes Diabetes Care. 2010;33(11):2327–2332
- 4. Mingrone G, et al Use of dicarboxylic acids in type 2 diabetes Br J Clin Pharmacol. 2013;75(3):671–676
- 5. Greco AV, et al The metabolic effect of dodecanedioic acid infusion in non-insulin-dependent diabetic patients Nutrition. 1998;14(4):351–357
- 6. Raguso CA, et al Dicarboxylic acids and glucose utilization in humans: effect of sebacate JPEN J Parenter Enteral Nutr. 1994;18(1):9–13
- 7. Salinari S, et al Dodecanedioic acid overcomes metabolic inflexibility in type 2 diabetic subjects Am J Physiol Endocrinol Metab. 2006;291(5):E1051–E1058
- 8. Membrez M, et al Six weeks’ sebacic acid supplementation improves fasting plasma glucose, HbA1c and glucose tolerance in db/db mice Diabetes Obes Metab. 2010;12(12):1120–1126
- 9. Malaisse WJ, et al Oxidation of D-[U-(14)C] glucose and [1,12-(14)C] dodecanedioic acid by pancreatic islets from Goto-Kakizaki rats Horm Metab Res. 2001;33(8):463–466
- 10. Jaacks LM, et al The obesity transition: stages of the global epidemic Lancet Diabetes Endocrinol. 2019;7(3):231–240
- 11. Ferrannini G, et al Genetic backgrounds determine brown remodeling of white fat in rodents Mol Metab. 2016;5(10):948–958
- 12. National Diabetes Statistics Report 2020. https://www.cdc.gov/diabetes/pdfs/data/statistics/national-diabetes-statistics-report.pdf Accessed May 9, 2024
- 13. Akiyama T, et al High-fat hypercaloric diet induces obesity, glucose intolerance and hyperlipidemia in normal adult male Wistar rat Diabetes Res Clin Pract. 1996;31(1-3):27–35
- 14. Eslam M, et al MAFLD: A consensus-driven proposed nomenclature for metabolic associated fatty liver disease Gastroenterology. 2020;158(7):1999–2014
- 15. Younossi ZM, et al The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): a systematic review Hepatology. 2023;77(4):1335–1347
- 16. Verrastro O, et al Bariatric-metabolic surgery versus lifestyle intervention plus best medical care in non-alcoholic steatohepatitis (BRAVES): a multicentre, open-label, randomised trial Lancet. 2023;401(10390):1786–1797
- 17. Harrison SA, et al A phase 3, randomized, controlled trial of resmetirom in NASH with liver fibrosis N Engl J Med. 2024;390(6):497–509
Conflict of interest: LCG is scientific advisor of Jemyll Ltd. GM reports consulting fees from Novo Nordisk, Eli Lilly, Boehringer Ingelheim, Fractyl Inc, and Recor Inc. She is also scientific advisor of Metadeq, Keyron, GHP Scientific, and Jemyll Ltds.
Copyright: © 2024, Castagneto-Gissey et al. This is an open access article published under the terms of the Creative Commons Attribution 4.0 International License.
Reference information: J Clin Invest. 2024;134(12):e181978. https://doi.org/10.1172/JCI181978.
See the related article at Dietary dicarboxylic acids provide a nonstorable alternative fat source that protects mice against obesity.