Schistosomiasis is a neglected parasitic infection affecting 250 million people worldwide [1,2]. Schistosoma mansoni infection, prevalent in sub-Saharan Africa [], is associated with gastrointestinal and liver injury through direct and immune-mediated mechanisms []. However, less attention has focused on genital tract effects of S. mansoni.
Population-based and autopsy studies have established that S. mansoni eggs can be found in female genital tissue, particularly the cervix and vagina []. Several studies have reported a possible increased susceptibility to human immunodeficiency virus (HIV), or increased HIV viral entry into cervical cells, among women with S. mansoni infection, implying that cervical immune alterations may be associated with infection []. Furthermore, treatment of S. mansoni reduced entry of HIV into cervical CD4+ T cells []. How S. mansoni alters the cervical immune compartment and mechanisms by which S. mansoni could impair genital mucosal antiviral immunity have been minimally investigated. Understanding this interaction could provide insight into immune effects of S. mansoni in the genital tract and help explain disparities in viral infections in women in countries where schistosomiasis is endemic [].
Available limited human data suggest that schistosome infection could alter mucosal immune cell populations in the female genital tract, but these have not been well characterized in S. mansoni infection [14,15]. In mouse models, infection with S. mansoni or other helminths decreased subsequent mucosal cytotoxic CD8+ T-cell responses to viruses in gastrointestinal mucosa [16,17]. Further murine studies indicate that alternatively activated type 2 (immunomodulatory) macrophages play a major role in the tissue immune response to S. mansoni eggs by promoting tissue granuloma formation and fibrosis, while concomitantly decreasing proinflammatory type 1 antiviral responses []. Notably, mouse models have focused on gastrointestinal and not genital mucosa.
To characterize genital immune cell composition in human S. mansoni infection, we studied mononuclear cells from cervical brushings of women with and without S. mansoni infection in Tanzania. We hypothesized that women with S. mansoni infection would have altered cervical immune cell frequencies compared to uninfected women, and we particularly focused on immune cells that may play a role in the antiviral mucosal immune response including natural killer (NK) cells, NK T cells, cytotoxic CD8+ cells, and monocytes. Our goal was to identify alterations in cervical immune cells and potential therapeutic targets that could restore genital mucosal abnormalities that persist despite antiparasitic treatment [].
METHODS
Overview
We studied baseline cervical samples from women aged 18–50 years participating in a longitudinal study in Tanzania in 2017. The study, conducted in rural communities near Lake Victoria among women who had limited access to clean water, has been previously described []. In brief, women provided written informed consent and underwent voluntary counseling and testing for HIV and screening for schistosome infection via egg detection in urine and stool, and circulating anodic antigen (CAA) testing in serum. Women who were HIV uninfected and had confirmed S. mansoni infection with stool eggs and serum CAA ≥30 pg/mL were invited to participate. A similar number of women without schistosome infection (egg negative and serum CAA <30 pg/mL) were also invited. In these communities, the prevalence of S. mansoni infection among adult women ranges from approximately 35% to 80%, while the prevalence of Schistosoma haematobium is approximately 2% []. See the Supplementary Appendix for details.
Cervical Mononuclear Cell Collection
Cervical mononuclear cells were collected during gynecologic examination using 2 endocervical cytobrushes and 1 Ayer spatula rotated 360° around the face of the os [26,27]. Cells were placed in phosphate-buffered saline (PBS), transported to Tanzania's National Institute for Medical Research laboratory within 4 hours of collection, vortexed, and the Ayer spatula discarded. Cytobrushes were scraped into a 15-mL Falcon tube using a 25-mL pipette while discharging PBS, as previously described with minor modifications []. Cells were washed, resuspended in 20% dimethyl sulfoxide, and cooled to −80°C using CoolCell (Corning), then to −156°C. Samples were shipped to Weill Cornell in New York at −150°C and stored in liquid nitrogen.
Flow Cytometry
Cells were washed twice in media containing 20% fetal bovine serum in RPMI and resuspended in 1 mL of the same media. Cells were then washed with Stain Buffer (BD Pharmigen) and incubated with Fc Receptor Blocking Solution (Biolegend) for 5 minutes. Cells were stained for 30 minutes in the dark with the following antibodies: CD14 PE, CD8 perCP-Cy5.5, CD56 PE-Dazzle 594, CD19 APC, CD4 PE-Cy7, CD45 Alexa700, CD3 BV711, and DAPI (all Biolegend). Compensation controls were prepared using UltraComp eBeads (Thermo Fisher). Cells and beads were then washed and resuspended in Stain Buffer (BD Pharmigen).
Flow cytometry was performed using a BD LSR Fortessa flow cytometer, equipped with 355-, 405-, 488-, 561-, and 640-nm lasers, in Weill Cornell's Flow Cytometry Core. Cleaning and gating were performed using FlowJo (BD). Fluorescence Minus One controls using healthy-donor peripheral blood mononuclear cells were created to obtain reliable gating parameters. A gating strategy was used to define 11 cell types, all derived from a parent DAPI−/CD45+ gate (Supplementary Table 1). A subset of samples underwent additional staining and gating that included markers for neutrophils, eosinophils, and basophils (Supplementary Table 2).
Statistical Analyses
Analyses were performed using Stata 15.1 (StataCorp). We summarized continuous variables with median and interquartile range [IQR] and categorical variables with numbers and percentages. We used rank-sum tests to compare median values between participants, including percentages of cell types out of total CD45+ cells. We compared categorical variables using Fisher exact or χ2 tests where appropriate.
Patient Consent Statement
All participants provided written informed consent, with ethical approvals obtained in New York and Tanzania (Supplementary Appendix).
RESULTS
Participant Characteristics
Sixty-four cervical samples were available for testing. Of these, 31 had <1000 viable CD45+ cells and were excluded. The remaining 33 samples included in the analysis had a median CD45+ cell count of 6067 total cells [IQR, 3130–12 586]. Among these 33 women, 13 (39%) had S. mansoni infection. Most factors were similar between those with S. mansoni and those without (Table 1). Women with S. mansoni infection had a median serum CAA of 1527.1 [IQR, 635.7–4680] pg/mL and 19.2 [IQR, 9.6–43.2] eggs per gram of stool. No differences were noted between the 31 women excluded for insufficient CD45+ cells and the 33 who were included (Supplementary Table 3).
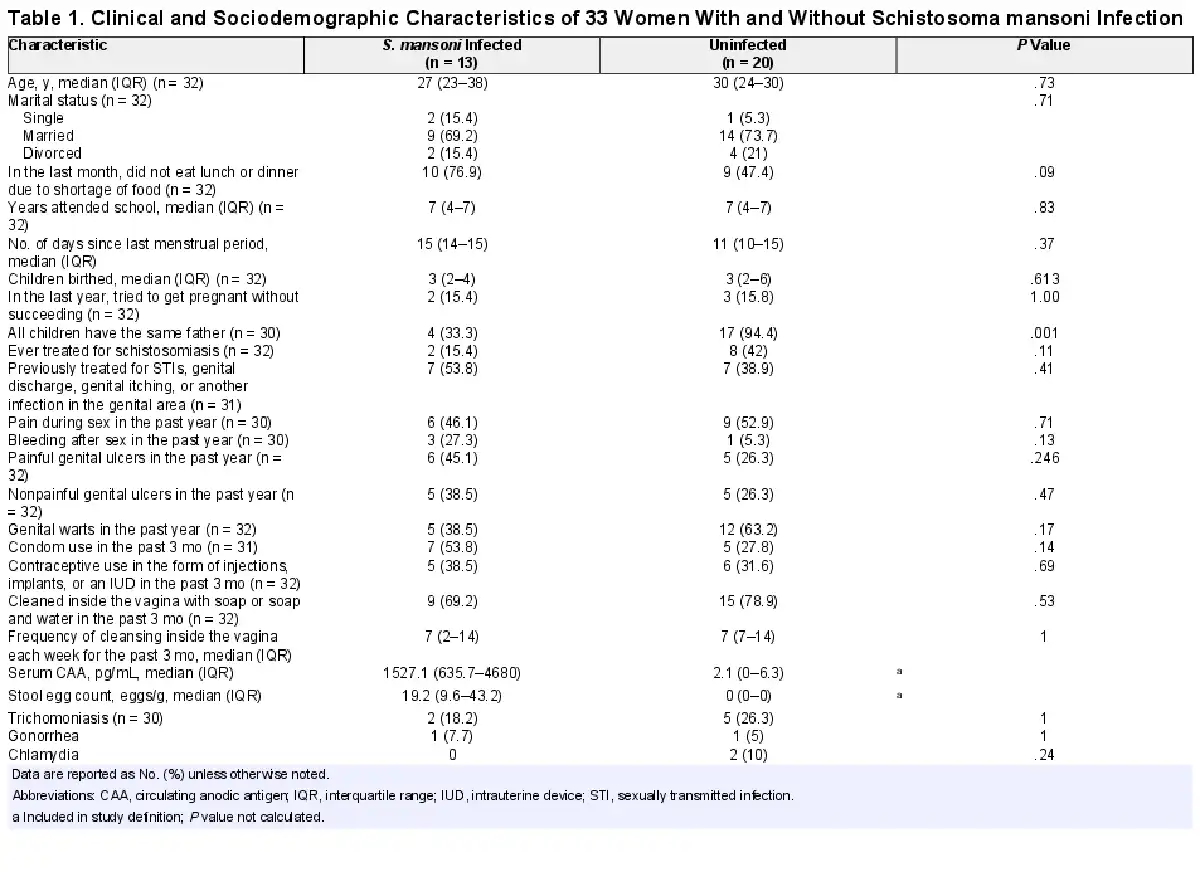
Cervical Mononuclear Cell Frequencies
Natural killer T cells (CD56+CD3+) were less frequent among women with S. mansoni infection than those without (median 1.7% [IQR, 1%–2.8%] vs 2.9% [2.0%–6.6%], P = .017; Figure 1). A similar trend was observed among NK cells (CD56+CD3−: 3.7% [2.6%–6.5%] vs 6.5% [4.5%–9.9%], P = .065). Women with S. mansoni also had higher frequencies of CD14+ monocytes (13.3% [9.8%–29%] vs 8.1% [3.7%–18.4%], P = .03). No differences in other cell type frequencies, including cytotoxic CD8+ T cells or helper CD4+ T cells, were noted.
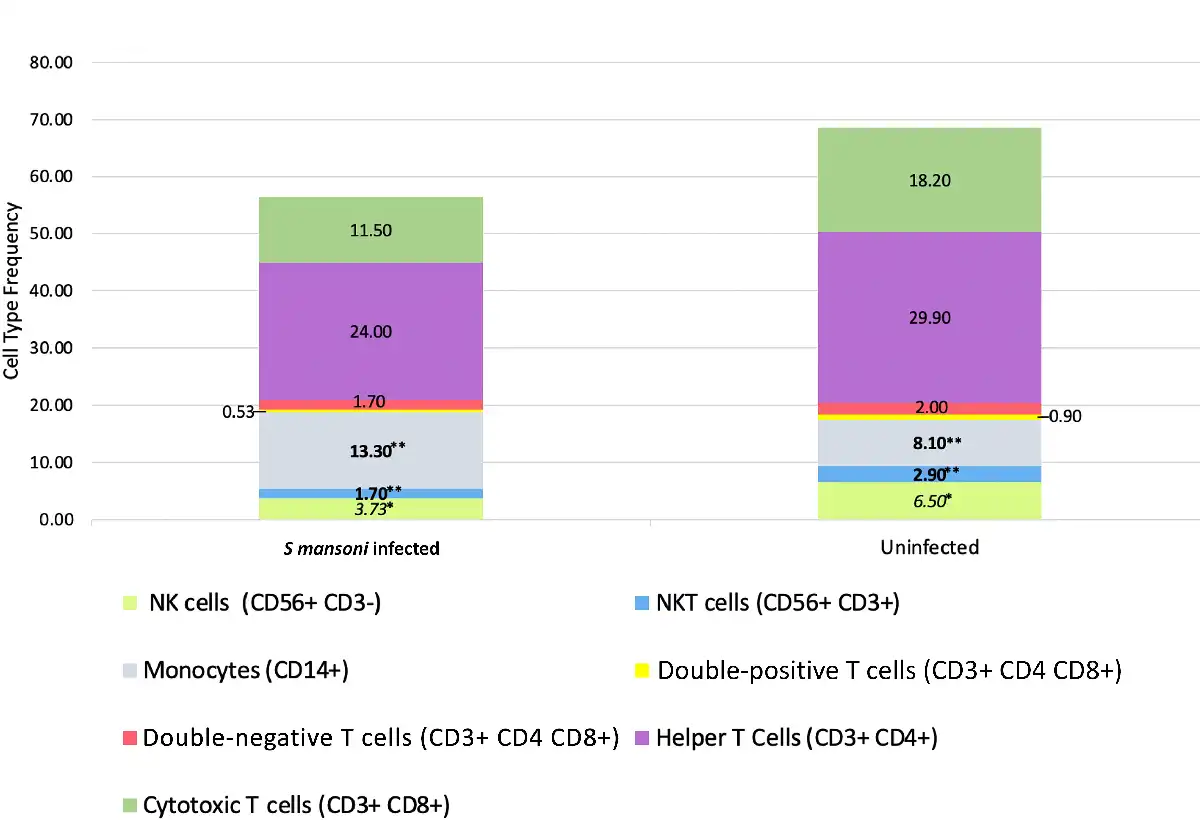
Figure 1
Cervical mononuclear cell types in women with and without Schistosoma mansoni infection. Bar graph containing median frequencies of cervical mononuclear cell types (as percentage of CD45+DAPI− cells) in women with (n = 13) and without (n = 20) S. mansoni infection. Cell types quantified include monocytes, T-cell subsets, natural killer (NK) cells, and natural killer T (NKT) cells. Differences between frequencies: *P < .10, **P < .05.
A subset of 8 women with S. mansoni infection and 14 without had additional neutrophil, eosinophil, and basophil markers included. Frequencies of these cells did not differ by S. mansoni infection status (Supplementary Figure 1).
DISCUSSION
We report that the human cervical immune compartment is altered in S. mansoni infection. These data align with prior studies in mice and humans demonstrating that helminth infections could cause mucosal immune dysregulation, which in S. mansoni infection was associated with increased HIV viral entry [10,16,17,28]. Specifically, we found lower NK T cells, a trend toward lower NK cells, and increased CD14+ monocytes in S. mansoni infection. We note our focus on S. mansoni infection, not classically regarded as having urogenital effects, in genital mucosal immune populations.
Our findings point in the same direction as observations of increased viral infection and impaired genital antiviral immunity in women with S. mansoni. Lower NKT and NK cell frequencies could reduce mucosal antiviral immunity. Given that NKT cell proliferation is often accompanied and promoted by interferon secretion [29,30], these findings are in agreement with previously reported dysregulated interferon secretion from cervical cells of Kenyan women with S. mansoni []. Similarly, increased CD14+ monocytes, particularly if they represent an increase in the type 2 to type 1 monocyte ratio, would also be expected to decrease mucosal antiviral immunity [16,18]. Together with observations of schistosomes impairing host immunity to a variety of viral infections [], our data could indicate clinically meaningful pathways by which schistosome infection may impair control of viral infections in the genital mucosa.
Our data supplement only 2 other studies of cervical mucosal immune cells in schistosome infections. In the eloquent Kenyan study described above, investigators reported no change in CD4+ cervical cells pre- and posttreatment for S. mansoni, but did not describe other cervical mononuclear populations. A study of South African women with S. haematobium infection reported decreased frequency of cervical CD14+ cells following praziquantel treatment, which could be consistent with our findings of increased CD14+ cells at baseline []. This South African study reported that <2% of cervical mononuclear cells were CD14+, contrasting with findings from our study and others that CD14+ cells constitute 10%–30% of mononuclear cells [15,26,31]. We confirm findings from these studies regarding pretreatment CD4+ and CD14+ frequencies, and add data on lower NK and NKT cell frequencies in S. mansoni infection.
This study had strengths and limitations. It is the first to quantify the proportions of genital immune cells broadly in relation to schistosome infection. As in many cervical cell isolation studies, approximately one-half of study samples had insufficient cell numbers for stringent analysis and were excluded, limiting sample size. Furthermore, we were limited to few cell surface markers given flow cytometry capabilities; future studies incorporating more advanced techniques such as spectral flow or mass cytometry could further characterize CD14+ monocytes and quantify other cells of interest such as regulatory T and dendritic cells.
Future longitudinal studies to quantify effects of S. mansoni on genital mucosal immune cell populations, both before and after treatment, are needed. Such analysis will identify persistent alterations and point toward targeted therapies, potentially including host-directed immunomodulatory agents, that could be combined with praziquantel to promote mucosal healing in women suffering from genital tract sequelae of schistosome infections.
References
- 1. Lo NC, Bezerra FSM, Colley DG, et al Review of 2022 WHO guidelines on the control and elimination of schistosomiasis. Lancet Infect Dis2022; 22:e327–35.
- 2.
- 3.
- 4. Verjee MA. Schistosomiasis: still a cause of significant morbidity and mortality. Res Rep Trop Med2019; 10:153–63.
- 5. Poggensee G, Krantz I, Kiwelu I, Diedrich T, Feldmeier H. Presence of Schistosoma mansoni eggs in the cervix uteri of women in Mwanga District, Tanzania. Trans R Soc Trop Med Hyg2001; 95:299–300.
- 6. Gelfand M, Ross MD, Blair DM, Weber MC. Distribution and extent of schistosomiasis in female pelvic organs, with special reference to the genital tract, as determined at autopsy. Am J Trop Med Hyg1971; 20:846–9.
- 7. Cheever AW, Kamel IA, Elwi AM, Mosimann JE, Danner R. Schistosoma mansoni and S. haematobium infections in Egypt. II. Quantitative parasitological findings at necropsy. Am J Trop Med Hyg1977; 26:702–16.
- 8. Gelfand M, Ross WFII. The distribution of schistosome ova in the genito-urinary tract in subjects of bilharziasis. Trans R Soc Trop Med Hyg1953; 47:218–20.
- 9. Edington GM, Nwabuebo I, Junaid TA. The pathology of schistosomiasis in Ibadan, Nigeria with special reference to the appendix, brain, pancreas, and genital organs. Trans R Soc Trop Med Hyg1975; 69:153–6.
- 10. Yegorov S, Joag V, Galiwango RM, et al Schistosoma mansoni treatment reduces HIV entry into cervical CD4+ T cells and induces IFN-I pathways. Nat Commun2019; 10:2296.
- 11. Wall KM, Kilembe W, Vwalika B, et al Schistosomiasis is associated with incident HIV transmission and death in Zambia. PLoS Negl Trop Dis2018; 12:e0006902.
- 12. Downs JA, Dupnik KM, van Dam GJ, et al Effects of schistosomiasis on susceptibility to HIV-1 infection and HIV-1 viral load at HIV-1 seroconversion: a nested case-control study. PLoS Negl Trop Dis 2017; 11:e0005968.
- 13. Bullington BW, Klemperer K, Mages K, et al Effects of schistosomes on host anti-viral immune response and the acquisition, virulence, and prevention of viral infections: a systematic review. PLoS Pathog2021; 17:e1009555.
- 14. Feldmeier H, Daccal RC, Martins MJ, Soares V, Martins R. Genital manifestations of schistosomiasis mansoni in women: important but neglected. Mem Inst Oswaldo Cruz1998; 93:127–33.
- 15. Kleppa E, Ramsuran V, Zulu S, et al Effect of female genital schistosomiasis and anti-schistosomal treatment on monocytes, CD4+ T-cells and CCR5 expression in the female genital tract. PLoS One2014; 9:e98593.
- 16. Osborne LC, Monticelli LA, Nice TJ, et al Virus-helminth coinfection reveals a microbiota-independent mechanism of immunomodulation. Science2014; 345:578–82.
- 17. Actor JK, Shirai M, Kullberg MC, Buller RM, Sher A, Berzofsky JA. Helminth infection results in decreased virus-specific CD8+ cytotoxic T-cell and Th1 cytokine responses as well as delayed virus clearance. Proc Natl Acad Sci U S A1993; 90:948–52.
- 18. Reese TA, Wakeman BS, Choi HS, et al Coinfection. Helminth infection reactivates latent γ-herpesvirus via cytokine competition at a viral promoter. Science2014; 345:573–7.
- 19. McSorley HJ, Hewitson JP, Maizels RM. Immunomodulation by helminth parasites: defining mechanisms and mediators. Int J Parasitol2013; 43:301–10.
- 20. Herbert DR, Hölscher C, Mohrs M, et al Alternative macrophage activation is essential for survival during schistosomiasis and downmodulates T helper 1 responses and immunopathology. Immunity2004; 20:623–35.
- 21. Downs JA, Kabangila R, Verweij JJ, et al Detectable urogenital schistosome DNA and cervical abnormalities 6 months after single-dose praziquantel in women with Schistosoma haematobium infection. Trop Med Int Health 2013; 18:1090–96.
- 22.
- 23. Kapiga S, Hansen CH, Downs JA, et al The burden of HIV, syphilis, and schistosome infection and associated factors among adults in the fishing communities in northwestern Tanzania. Trop Med Int Health2021; 26:204–13.
- 24. Downs JA, Mguta C, Kaatano GM, et al Urogenital schistosomiasis in women of reproductive age in Tanzania's Lake Victoria region. Am J Trop Med Hyg2011; 84:364–9.
- 25. Downs JA, van Dam GJ, Changalucha JM, et al Association of schistosomiasis and HIV infection in Tanzania. Am J Trop Med Hyg2012; 87:868–73.
- 26. McKinnon LR, Hughes SM, De Rosa SC, et al Optimizing viable leukocyte sampling from the female genital tract for clinical trials: an international multi-site study. PLoS One2014; 9:e85675.
- 27. Juno JA, Boily-Larouche G, Lajoie J, Fowke KR. Collection, isolation, and flow cytometric analysis of human endocervical samples. J Vis Exp2014; 89:51906.
- 28. Chetty A, Darby MG, Vornewald PM, et al IL4RA-independent vaginal eosinopnil accumulation following helminth infection exacerbates epithelial ulcerative pathology of HSV-2 infection. Cell Host Microbe2021; 29:579–93.e5.
- 29. Ivashkiv LB. IFNγ: signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat Rev Immunol2018; 18:545–58.
- 30. Jabri B, Abadie V. IL-15 functions as a danger signal to regulate tissue-resident T cells and tissue destruction. Nat Rev Immunol2015; 15:771–83.
- 31. Trifonova RT, Lieberman J, van Baarle D. Distribution of immune cells in the human cervix and implications for HIV transmission. Am J Reprod Immunol2014; 71:252–64.